Трематоды это: Трематоды – это… Что такое Трематоды?
Распознать паразитофауну
Архангельские ученые предложили метод выявления и идентификации трематод в пресноводных улитках без проведения визуального обследования, что поможет оценить паразитарную обстановку в окружающей среде
Брюхоногие моллюски семейства Lymnaeidae, или прудовики, которые обитают повсеместно и, в том числе, на территории Арктики, могут выступать в качестве промежуточного хозяина для определенных видов паразитов. В ходе миграции животных, таких, например, как рыбы и птицы, паразиты распространяются из одного региона в другой. Чаще всего, такими представителями паразитофауны становятся трематоды – класс плоских паразитических червей, насчитывающий несколько тысяч видов. Они являются источником опасных заболеваний у животных и человека. Поэтому новые методы определения паразитов нужны как для сохранения биоразнообразия в целом, так и для обеспечения экологической безопасности арктических территорий.
Специалисты ФГБУН ФИЦКИА УрО РАН (г.
Александр Васильевич Кондаков – кандидат технических наук
Сотрудники Института биогеографии и генетических ресурсов Федерального исследовательского центра комплексного изучения Арктики им. акад. Н.П. Лаверова УрО РАН (г. Архангельск) – кандидат технических наук, ведущий научный сотрудник лаборатории молекулярной экологии и биогеографии Александр Васильевич Кондаков и кандидат биологических наук, ведущий научный сотрудник Российского музея центров биологического разнообразия Ольга Владимировна Аксёнова – рассказали о результатах исследований трематод в организме пресноводных брюхоногих моллюсков семейства Lymnaeidae и о предложенном авторским коллективом эффективном молекулярно-генетическом методе определения видового состава внутренних паразитов трематод позвоночных и беспозвоночных животных.
Ольга Владимировна Аксёнова – кандидат биологических наук
По мнению Александра Кондакова, «очень часто люди вспоминают про паразитов, когда каким-либо образом заражаются трематодами или обнаруживают их, например, в рыбе, дичи или у сельскохозяйственных животных. Но даже из школьной программы по биологии известно, что трематоды имеют сложный жизненный цикл со сменой промежуточных хозяев, одними из которых чаще всего выступают пресноводные брюхоногие моллюски, которых мы называем прудовиками или в народе просто улитками. И изначально наши исследования были направлены в целом на выявление трематод в прудовиках молекулярными методами. Но в одной из работ мы объединили полученные данные по этим животным в Арктике, т.к. в настоящее время этот вопрос действительно актуален».
«Все мы неоднократно слышали про климатические изменения, которые наиболее ярко проявляются именно в Арктике. Происходящие здесь изменения могут способствовать активизации таких процессов, как увеличение численности паразитов и их переносчиков, частичная или полная замена паразитофауны хозяев, а также проникновение паразитов на новые территории. Многие исследователи все чаще отмечают появление ранее не встречающихся здесь видов животных и растений. Не являются исключением моллюски и развивающиеся в них трематоды, которые получают возможность пройти полный жизненный цикл в наших северных водоемах, а значит в итоге попасть в организм человека или животного. В связи с этим обнаружение и определение видового состава паразитов на арктических территориях имеет большое значение для оценки существующей паразитарной обстановки и прогнозирования её изменений с целью предотвращения заражений», – дополняет ответ коллеги Ольга Аксёнова.
Многие исследователи все чаще отмечают появление ранее не встречающихся здесь видов животных и растений. Не являются исключением моллюски и развивающиеся в них трематоды, которые получают возможность пройти полный жизненный цикл в наших северных водоемах, а значит в итоге попасть в организм человека или животного. В связи с этим обнаружение и определение видового состава паразитов на арктических территориях имеет большое значение для оценки существующей паразитарной обстановки и прогнозирования её изменений с целью предотвращения заражений», – дополняет ответ коллеги Ольга Аксёнова.
Занимаясь исследованием прудовиков (в настоящее время проанализировано порядка 700 образцов), специалисты обнаружили в их органах и тканях наличие паразитов. Процент инфицирования моллюсков партенитами трематод в среднем составил около 12%, то есть практически каждая десятая особь из исследованных была заражена.
В большинстве своем, как заметил Александр Кондаков, «трематод изучают уже в половозрелом возрасте, когда обнаруживают их в животных.
Материал для исследования сотрудниками ФГБУН ФИЦКИА УрО РАН был собран в ходе многочисленных экспедиций в разных уголках России: от Мурманской области до Чукотки, а также за рубежом (например, в Китае, Корее и США). Исследования были поддержаны несколькими грантами, в т.ч. грантом Президента РФ № МК-1720.2019.4 «Изучение таксономического и генетического разнообразия брюхоногих моллюсков семейства Lymnaeidae как промежуточных хозяев трематод с применением современных молекулярно-генетических и биогеографических подходов» и продолжаются в рамках проекта РНФ № 21-74-10155 «Пресноводные моллюски России: интегративная таксономия, биогеография, экология и паразитофауна».
Район исследований: красными точками отмечены места сборов образцов прудовиков в ходе экспедиционных исследований
Авторы: М.Ю. Гофаров, А.В. Кондаков
В каком регионе/районе больше всего прудовиков, пораженных трематодами и почему?
«Считается, что трематодозы – заболевания, вызванные трематодами и наиболее распространены в тропических странах, а на территории России встречаются в Сибири и на Дальнем Востоке. Но следует отметить, что не всегда в этом виноваты именно прудовики. Здесь в качестве промежуточных хозяев выступают и другие семейства брюхоногих моллюсков, и связано это больше с природными особенностями регионов, которые способствуют большему разнообразию промежуточных хозяев и благоприятствуют развитию трематод в них. Что касается прудовиков, то пока сложно выделить регионы, где они больше заражены, а где меньше, т.к. исследование начато относительно недавно, но распространены они практически во всех пресных водоемах»,
По замечанию Александра Кондакова, «нет таких регионов, где наблюдается высокое инфицирование прудовиков трематодами. В целом процент инфицированных моллюсков везде около 12%. Исключением являются искусственно подогретые водоемы (каналы-охладители ТЭЦ), в которых моллюсков, инфицированных трематодами, нами обнаружено не было. Возможно, это связано с высокими температурами в этих водоемах, которые трематоды не способны переносить».
Прудовик обыкновенный (Lymnaea stagnalis) в одном из водоемов на юге полуострова Таймыр
Автор фото О.В. Аксёнова
Как прудовики переносят в своем организме наличие трематод: болеют, умирают?
«По этому вопросу есть несколько интересных исследований. В одном из которых говорится о том, что при инфицировании моллюска трематодами на ранних этапах развития замедляется его рост и в результате изменяется форма их раковин. В другом же исследовании определено, что трематоды осуществляют химическую кастрацию прудовика, то есть прудовик не может размножаться, продолжая жить и питаться, но при этом не тратит энергию на размножение. Но нужно четко понимать, что не в интересах трематод сильно угнетать промежуточного хозяина, так как они питаются за счет него», – ответил Александр Кондаков.
Но нужно четко понимать, что не в интересах трематод сильно угнетать промежуточного хозяина, так как они питаются за счет него», – ответил Александр Кондаков.
«Изначально свободноплавающая личинка трематод мирацидий находит моллюска в воде и проникает в его тело, а затем начинает там развиваться и размножаться. Происходит это, как правило, в половой железе или пищеварительной системе моллюска. В конце концов они достигают большой численности и заполняют внутреннюю полость тела моллюска, и это однозначно его угнетает, ослабляет иммунитет, влияет на его активность, поведение и продолжительность жизни. Есть примеры, когда зараженные прудовики достигают невероятно крупных размеров, т.е. трематоды ускоряют его развитие до наступления половозрелости, а затем начинают сами активно в нем развиваться», –
Почему трематоды выбирают моллюсков семейства Lymnaeidae?
«На самом деле трематоды используют в качестве промежуточных хозяев и другие семейства брюхоногих моллюсков. Сказать наверняка, почему они в основном выбирают в качестве промежуточных хозяев именно моллюсков семейства Lymnaeidae, сложно. Просто в нашем исследовании акцент был сделан именно на это семейство, как наиболее распространенное, но недостаточно изученное. Скорее всего это связано с коэволюционными процессами, которые привели к тому, что моллюски оказались оптимальным организмом для прохождения отдельных стадий развития трематод», – прокомментировал Александр Кондаков.
Сказать наверняка, почему они в основном выбирают в качестве промежуточных хозяев именно моллюсков семейства Lymnaeidae, сложно. Просто в нашем исследовании акцент был сделан именно на это семейство, как наиболее распространенное, но недостаточно изученное. Скорее всего это связано с коэволюционными процессами, которые привели к тому, что моллюски оказались оптимальным организмом для прохождения отдельных стадий развития трематод», – прокомментировал Александр Кондаков.
Где преимущественно трематоды обнаружены: на мантии или на внутренних органах прудовиков?
«Трематоды у зараженных моллюсков обычно локализуются во внутренних органах, в пищеварительной системе (гепатопанкреасе), в половой железе (гонаде) и в мышечной ткани (ноге) моллюска. Нередко их можно обнаружить и под мантией, но обычно моллюска нужно вскрыть, чтобы понять, заражен он или нет», – уточнила Ольга Аксенова.
Чем трематоды отличаются от остальных червей и по каким морфологическим признакам?
«Трематоды, или еще их называют сосальщики – это целый класс паразитических плоских червей.
Схема жизненного цикла трематод рода Diplostomum
Источник: Определитель паразитов пресноводных рыб СССР. – Изд-во АН СССР, 1962
Если говорить о взрослой стадии (марите), которую мы обычно можем встретить в мясе или внутренних органах позвоночных животных, в т. ч. и человека, то, соответственно, форма тела у них уплощенная и может быть разной в зависимости от семейства: грушевидная, листовидная или нитевидная. Как правило, на теле у них есть ротовая и брюшная присоски, но опять же не у всех. Размеры их могут варьировать от нескольких миллиметров до нескольких сантиметров. На стадии свободноплавающей личинки мирацидия до попадания в промежуточного хозяина – например, их тело покрыто чувствительными ворсинками, на стадии церкария – после развития в моллюске, у них вырастает хвостик, который тоже у разных видов может быть нитевидным или раздваиваться на конце, а на стадии метацеркария, когда трематода развивается во втором промежуточном хозяине – хвостик отпадает. Т.е. довольно сложно выделить какие-то четкие признаки, благодаря которым неспециалист бы увидел и с легкостью сказал, что это именно трематода», – детально информировала Ольга Аксёнова.
ч. и человека, то, соответственно, форма тела у них уплощенная и может быть разной в зависимости от семейства: грушевидная, листовидная или нитевидная. Как правило, на теле у них есть ротовая и брюшная присоски, но опять же не у всех. Размеры их могут варьировать от нескольких миллиметров до нескольких сантиметров. На стадии свободноплавающей личинки мирацидия до попадания в промежуточного хозяина – например, их тело покрыто чувствительными ворсинками, на стадии церкария – после развития в моллюске, у них вырастает хвостик, который тоже у разных видов может быть нитевидным или раздваиваться на конце, а на стадии метацеркария, когда трематода развивается во втором промежуточном хозяине – хвостик отпадает. Т.е. довольно сложно выделить какие-то четкие признаки, благодаря которым неспециалист бы увидел и с легкостью сказал, что это именно трематода», – детально информировала Ольга Аксёнова.
По словам ученых, наличие трематод в моллюсках свидетельствует о неблагополучной паразитарной обстановке в водоеме или в регионе в целом. Т.е. если трематоды есть в моллюсках, то скорее всего есть и окончательный хозяин, который будет заражен рано или поздно. Это могут быть рыбы, птицы, домашний скот или человек. Чаще всего трематоды расселяются за счет птиц, которые осуществляют перенос паразитов на большие расстояния.
Т.е. если трематоды есть в моллюсках, то скорее всего есть и окончательный хозяин, который будет заражен рано или поздно. Это могут быть рыбы, птицы, домашний скот или человек. Чаще всего трематоды расселяются за счет птиц, которые осуществляют перенос паразитов на большие расстояния.
Каков риск заражений ими человека и животных?
«Риск заражения трематодами всегда есть. Для человека источником заражения могут быть сырое мясо или рыба, или можно просто искупаться в водоеме, где есть трематоды. Есть виды трематод, которые способны проникать через кожу, или человек или животное может просто заглотить с водой или растениями их личинки», – отвечает Ольга Аксёнова.
«Есть интересная статистика, отражающая прямую зависимость увеличения потребления сырой рыбы в еду в виде суши и количеством паразитических заболеваний у человека. Поэтому потреблять сырую рыбу и мясо в пищу небезопасно. А животные в меньшей степени защищены от паразитов, чем люди, так как часто питаются растениями и пьют воду непосредственно из водоемов», – сообщил Александр Кондаков.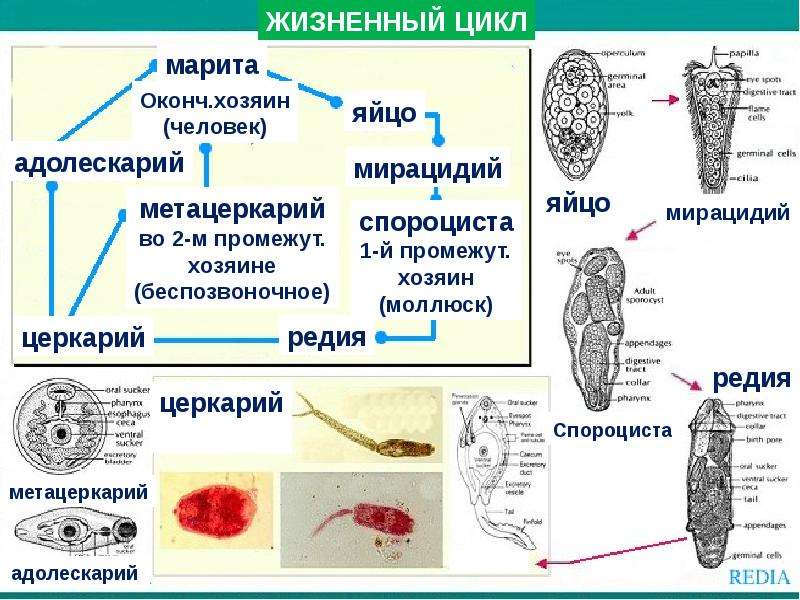
В современной исследовательской практике для идентификации трематод применяют различные молекулярно-генетические методы. Сотрудники ФГБУН ФИЦКИА УрО РАН разработали и запатентовали оригинальный метод для выявления и идентификации трематод в моллюске без проведения визуального обследования. Таким образом за период исследований было обнаружено 24 вида трематод, относящихся к 12 родам.
Как подчеркивает Александр Кондаков, «методов, которые позволяют определить широкий спектр трематод одновременно, как таковых нет. Разработанный нами метод позволяет выявлять практически всех известных трематод в прудовиках и осуществлять их идентификацию. Это и является преимуществом нашего метода. Он дает возможность на ранних этапах развития трематод независимо от стадии определить их присутствие и видовой состав в пресных водоемах посредством молекулярно-генетического анализа моллюсков, которые выступают для них промежуточными хозяевами».
Алгоритм выявления и идентификации и трематод у моллюсков семейства Lymnaeidae
Авторы: А. В. Кондаков, И.С. Хребтова
В. Кондаков, И.С. Хребтова
В чем состоит ваш подход и как вы определяете принадлежность к различным биологическим видам?
«Суть метода заключается в следующем: мы берем образцы моллюсков из водоема, затем в лаборатории выделяем из них ДНК и проводим амплификацию (увеличение числа копий ДНК) с помощью полимеразной цепной реакции (ПЦР). В результате при анализе продуктов, полученных в ходе реакции, мы уже можем увидеть на электрофореграмме заражен моллюск или нет. Грубо говоря, если видим одну полосу на рисунке, то моллюск не заражен, если две и более полос, то моллюск заражен. Если хотим определить вид моллюска и/или трематод, то далее подготавливаем продукт и проводим секвенирование, т.е. прочитав полученные фрагменты ДНК выбранных нами маркерных генов и сопоставив их с имеющейся в международной базе данных «GenBank» информацией, в результате мы узнаем видовую принадлежность интересующих нас образцов», – раскрыл детали авторского подхода Александр Кондаков.
Электрофореграмма с результатами амплификации генетического маркера рибосомной ДНК ITS2
Автор: И.С. Хребтова
Чем подтверждается надежность и точность вашего метода?
«Надежность и точность нашего метода подтверждается полученными результатами. Полученные нуклеотидные последовательности ДНК как штрих-код позволяют практически безошибочно определить вид, если информация о нем есть в базе или понять, что мы имеем дело с новым, еще малоизученным видом. Такое тоже бывает», – аргументировал ученый.
Где важны полученные молекулярные данные по изучению популяций прудовиков с точки зрения паразитологии? Как эти сведения способствуют пониманию фактического распределения определенных видов трематод?
«Полученные молекулярные данные по прудовикам позволяют определить видовой состав моллюсков в исследуемом регионе или отдельном водоеме. Проанализировав накопленные данные, мы можем реконструировать пути их расселения, определить особенности эволюции данной группы, уточнить их таксономический статус. Идентификация видов моллюсков позволяет определить наличие или отсутствие специфичности трематод по отношению к моллюскам-хозяевам, что важно при изучении коэволюции данных групп или при организации санитарных мероприятий. Данные о видовом составе трематод, спектре их промежуточных хозяев и их распространении позволят разрабатывать более эффективные меры борьбы с ними. Результаты исследования имеют важное практическое значение для оценки и мониторинга санитарно-эпидемиологического состояния поверхностных водоемов и предотвращения заражений человека и животных», – убеждены специалисты ФГБУН ФИЦКИА УрО РАН Ольга Аксёнова и Александр Кондаков.
Идентификация видов моллюсков позволяет определить наличие или отсутствие специфичности трематод по отношению к моллюскам-хозяевам, что важно при изучении коэволюции данных групп или при организации санитарных мероприятий. Данные о видовом составе трематод, спектре их промежуточных хозяев и их распространении позволят разрабатывать более эффективные меры борьбы с ними. Результаты исследования имеют важное практическое значение для оценки и мониторинга санитарно-эпидемиологического состояния поверхностных водоемов и предотвращения заражений человека и животных», – убеждены специалисты ФГБУН ФИЦКИА УрО РАН Ольга Аксёнова и Александр Кондаков.
Итак, своевременное и точное выявление паразитов у позвоночных и беспозвоночных животных при помощи надежной молекулярной идентификации позволит выработать меры, чтобы минимизировать риски заражения человека и животных.
ПАТОГЕННОЕ ВОЗДЕЙСТВИЕ ТРЕМАТОД И ИХ АССОЦИАЦИЙ НА ОРГАНИЗМ ХОЗЯИНА | Мкртчян
1. Абдуллаев Х. С. Формирование паразитарной системы в организме крупного рогатого скота и меры борьбы с паразитозами в Нечерноземной зоне Российской Федерации: автореф. дис. … д-ра вет. наук. – Иваново, 2007. – C. 26–27.
Абдуллаев Х. С. Формирование паразитарной системы в организме крупного рогатого скота и меры борьбы с паразитозами в Нечерноземной зоне Российской Федерации: автореф. дис. … д-ра вет. наук. – Иваново, 2007. – C. 26–27.
2. Васильев Ю. Г., Трошин Е. И., Любимов А. И. Ветеринарная клиническая гематология. – С.–П.: Лань, 2015. – С. 165–218.
3. Волков А. Х. Методы и средства борьбы с ассоциативными инвазионными болезнями крупного рогатого скота: дис… д-ра вет. наук. – Казань, 2000. – С. 69–130.
4. Джальчинова В. Б., Чистяков Г. М. Эозинофилы и их роль в патогенезе аллергических заболеваний. // Рос. вестник перинатологии и педиатрии. – 1999. – № 5. – С. 42–45.
5. Мазихова А. М. , Беккиева С. А., Юсупов А. О., Чиляев С. Ш. Влияние трематодозных инвазий на морфологический состав крови крупного рогатого скота // Вестник КрасГАУ. – 2009. – № 4. – С. 163–164.
, Беккиева С. А., Юсупов А. О., Чиляев С. Ш. Влияние трематодозных инвазий на морфологический состав крови крупного рогатого скота // Вестник КрасГАУ. – 2009. – № 4. – С. 163–164.
6. Петров Ю. Ф., Абдуллаев Х. С., Мухаммедов З. Р. и др. Динамика биохимических показателей при микстинвазии крупного рогатого скота. // Уч. Зап. КГАВМ. – Казань, 2006. – Т. 184. – С. 204–210.
7. Чухлебова О. М. Фармакологическая коррекция обменных процессов у крупного рогатого скота при гельминтозах и антигельминтной терапии: автореф. дис. … канд. вет. наук. – Воронеж, 2012. – С. 11–15.
8. Bush R. K., Geller M., Busse W. W. Response to corticosteroids in the hypereosinophilic syndrom. Arch. Intern. Med 1978; 138: 1244.
9. Buttervorth A. E., Sturrock R. F. Eosinophils as mediators antibodydependent damage to schistosomula. Nature 1975; 256: 727–729.
Buttervorth A. E., Sturrock R. F. Eosinophils as mediators antibodydependent damage to schistosomula. Nature 1975; 256: 727–729.
10. Prin L., Lefebvre P., Gruart V. Polinucleare eosinophile et recepteur glucocorticoide. Rev. Allergology. 1990; 30: 2: 83–85.
ТРЕМАТОДЫ | значение | Словарь иностранных слов
Словарь иностранных слов
трематод, ед. трематода, ы, ж. зоол.
Класс паразитических плоских червей, имеющих ротовую и брюшную присоски. | Взрослые особи трематод – паразиты позвоночных, личинки – паразиты беспозвоночных. ||Ср. НЕМАТОДЫ.
||Ср. НЕМАТОДЫ.
ТРЕМАТОДЫ
Что такое
ТРЕМАТОДЫ, ТРЕМАТОДЫ это, значение слова ТРЕМАТОДЫ, происхождение (этимология) ТРЕМАТОДЫ, синонимы к ТРЕМАТОДЫ, парадигма (формы слова) ТРЕМАТОДЫ в других словарях ► ТРЕМАТОДЫ – Т.Ф. Ефремова Новый словарь русского языка. Толково- словообразовательныйтремато́ды
мн.Название класса паразитических плоских червей, особенностью питания которых является всасывание пищи через ротовое отверстие; сосальщики.
► ТРЕМАТОДЫ – Современный толковый словарь изд. «Большая Советская Энциклопедия»(сосальщики) , класс паразитических плоских червей. Тело чаще листовидное, от нескольких мм до 1 м. Св. 5 тыс. (по другим данным, ок. 7 тыс.) видов. Развитие со сменой хозяев и чередованием поколений и сменой паразитических и свободноживущих фаз развития. Взрослые трематоды паразитируют в теле позвоночных животных и человека, вызывая трематодозы, партеногенетические поколения – большей частью в моллюсках. См., напр., Печеночная двуустка.
Взрослые трематоды паразитируют в теле позвоночных животных и человека, вызывая трематодозы, партеногенетические поколения – большей частью в моллюсках. См., напр., Печеночная двуустка.
сосальщики, отр. класса плоских червей, паразиты, живущие на коже, жабрах или во внутренних органах других животных.
(Источник: «Словарь иностранных слов, вошедших в состав русского языка». Павленков Ф., 1907)
► ТРЕМАТОДЫ – Словарь медицинских терминов(Trematoda; син. сосальщики дигенетические) класс типа плоских червей; представители Т. имеют нерасчлененное тело, снабженное присосками, и разветвленный, слепо заканчивающийся кишечник; окончательные хозяева – позвоночные животные, промежуточные хозяева – беспозвоночные, обычно моллюски; некоторые Т. являются паразитами человека.
Трематоды завербовали новых солдат в свою армию
Emlyn Resetarits, Mark Torchin & Ryan Hechinger / Biology Letters, 2020
Американские ученые показали, что в местах, где конкуренция трематод высока, численность касты солдат в одной улитке больше, чем в более спокойных районах. Остается неизвестным, как устанавливается баланс между мирной и воинствующей группами — отдельно в каждой колонии или на уровне популяции. Статья опубликована в журнале Biology Letters.
Остается неизвестным, как устанавливается баланс между мирной и воинствующей группами — отдельно в каждой колонии или на уровне популяции. Статья опубликована в журнале Biology Letters.
Защита от внешних угроз энергозатратна, и любой организм должен максимально эффективно распределять свои ресурсы в пользу размножения или обороны. При высоком уровне опасности нет смысла заботиться о самовоспроизведении, и приоритет должен отдаваться различным формам защиты. Это утверждение применимо и к индивидуальным организмам, и к общественным. Так, можно предположить, что животные, формирующие организованные колонии (пчелы и термиты, например), будут менять количество солдат в зависимости от уровня внешней угрозы. Однако данные, которые свидетельствуют в пользу этой гипотезы, скудны.
Пример социальных животных, удобный для изучения, — трематоды, которые паразитируют на калифорнийских улитках Cerithideopsis californica. После попадания внутрь моллюска один червь многократно клонирует себя и формирует колонию, которая распространяется по всему телу улитки. Однако паразиты из других колоний могут попробовать инфицировать того же хозяина, и тогда возникает необходимость обороняться. Для этого у трематод есть две касты: особи, задача которых — размножаться, и защитники. Защитники в несколько раз мельче репродуктивных червей и имеют более крупный рот. Известно, что численность армии трематод может меняться, однако условия, влияющие на это, изучены недостаточно тщательно.
Однако паразиты из других колоний могут попробовать инфицировать того же хозяина, и тогда возникает необходимость обороняться. Для этого у трематод есть две касты: особи, задача которых — размножаться, и защитники. Защитники в несколько раз мельче репродуктивных червей и имеют более крупный рот. Известно, что численность армии трематод может меняться, однако условия, влияющие на это, изучены недостаточно тщательно.
Группа ученых из Америки по руководством Эмилин Резетаритс (Emlyn Resetarits) из Университета Джорджии исследовала, зависит ли число воинов трематод от степени опасности их среды обитания. Для оценки уровня угрозы в той или иной географической точке они собрали и вскрыли 5520 улиток в 26 местах на западном побережье Северной Америки. Для каждой локации ученые подсчитали процент моллюсков, которые были заражены одним из шести видов трематод. На основе этого определили уровень опасности каждой точки для червей — чем больше улиток инфицировано, тем больше конкуренция за них и тем выше угроза для каждой отдельной колонии.
После этого ученые выбрали 168 колоний червей (одна улитка — одна колония), разрезали моллюсков на три части: мантию, среднюю часть и половые органы, где находится основная масса репродуктивных червей. В каждой трети подсчитали количество воинов и размножающихся особей. Исследователи предположили, что при возрастании угрозы количество защитников в мантии улитки, которая является линией обороны, будет увеличиваться.
В результате эксперимента авторы выяснили, что в зонах повышенной опасности число солдат, приходящихся на 1000 репродуктивных червей, существенно выше, чем в местах с меньшей конкуренцией. Это справедливо для пяти видов из шести, а отличие шестого вида от общей тенденции не подтверждается статистически.
Количество воинов, приходящихся на 1000 репродуктивных особей (вертикальная ось) в зависимости от уровня угрозы (горизонтальная ось) для шести видов трематод. Синим обозначены линии тренда, а красным – модель, работающая для всех трематод независимо от вида
Синим обозначены линии тренда, а красным – модель, работающая для всех трематод независимо от вида
Emlyn Resetarits, Mark Torchin & Ryan Hechinger / Biology Letters, 2020
Примечательно, что уровень угрозы не влиял на количество солдат, которые находились на «фронте», то есть в мантии улитки. По всей видимости, трематоды не используют стратегию вывода воинов на переднюю линию обороны, а лишь увеличивает общее количество защитников.
Понимание социобиологии трематод важно, так как они играют большую роль в своих экосистемах. Изучение этих паразитов может помочь узнать, как энергия распределяется в пищевых цепях, которые включают в себя червей, улиток, рыб и птиц. В дальнейшем авторы статьи планируют выяснить, самостоятельно ли каждая колония приспосабливается к уровню угрозы или же это происходит на уровне целой популяции.
Кроме трематод, на опасность реагируют и другие общественные животные. Например, при атаке хищника муравьи одной колонии собираются вместе и ведут себя, как единый организм, выбирая наиболее выгодную стратегию защиты.
Например, при атаке хищника муравьи одной колонии собираются вместе и ведут себя, как единый организм, выбирая наиболее выгодную стратегию защиты.
Алиса Бахарева
Трематоды (Trematoda) пресмыкающихся Среднего Поволжья Текст научной статьи по специальности «Биологические науки»
Известия Самарского научного центра Российской академии наук. 2011. Т. 13, № 5 УДК 576.895.122:598.1
ТРЕМАТОДЫ (TREMATODA) ПРЕСМЫКАЮЩИХСЯ СРЕДНЕГО ПОВОЛЖЬЯ
© 2011 А.А. Кириллов, Н.Ю. Кириллова
Институт экологии Волжского бассейна РАН, г. Тольятти
Поступила 20.03.2011
В работе обобщены сведения по видовому разнообразию трематод рептилий Среднего Поволжья. У пресмыкающихся Среднего Поволжья отмечается 21 вид трематод. Нами впервые для пресмыкающихся фауны России указывается трематода Neoglyphe sobolevi. Для Strigea sphaerula, larvae зарегистрирован новый хозяин – обыкновенная медянка. Разные виды рептилий отмечены в качестве случайных хозяев для 6 видов гельминтов. Приводятся описания и оригинальные рисунки 7 видов паразитов. Ключевые слова, трематоды, пресмыкающиеся, Среднее Поволжье.
У пресмыкающихся Среднего Поволжья отмечается 21 вид трематод. Нами впервые для пресмыкающихся фауны России указывается трематода Neoglyphe sobolevi. Для Strigea sphaerula, larvae зарегистрирован новый хозяин – обыкновенная медянка. Разные виды рептилий отмечены в качестве случайных хозяев для 6 видов гельминтов. Приводятся описания и оригинальные рисунки 7 видов паразитов. Ключевые слова, трематоды, пресмыкающиеся, Среднее Поволжье.
Волжский бассейн входит в ареалы 20 видов пресмыкающихся. Из них в Среднем Поволжье обитает 14 видов [3]. Гельминтологическому исследованию подвергнуто 9 видов рептилий: обыкновенный и водяной ужи, обыкновенная и степная гадюки, обыкновенная медянка, прыткая и живородящая ящерицы, ломкая веретеница, разноцветная ягцурка. Пресмыкающиеся региона в паразитологи-ческом отношении исследованы территориально крайне неравномерно. Несмотря на довольно широкое распространение рептилий сведений об их гельминтах вообще, и трематодах в частности, крайне мало.
Паразиты пресмыкающихся европейской части России и, в частности, Волжского бассейна изучены недостаточно. В Среднем Поволжье исследования гельминтов рептилий проводились мозаично, в основном в Татарстане, Нижегородской и Самарской областях. Сведения по трематодам пресмыкающихся Среднего Поволжья содержатся в работах ряда авторов [1, 4, 5, 9-15, 23-25, 32-34]. В то же время, нет сведений о фауне гельминтов рептилий Республики Марий Эл, Пензенской и Ульяновской областей.
Цель настоящей работы – на основании собственных исследований и литературного материала обобщить сведения по видовому разнообразию трематод пресмыкающихся Среднего Поволжья.
При этом нами не учитывались виды паразитов, находки которых у рептилий региона вызывает сомнение. К таким фактам относится находка у обыкновенного ужа в Татарстане Astiotrema reniferum (Looss, 1898) и А. odheneri Bhalerao, 1936 – облигатных паразитов дальневосточной черепахи [1].
odheneri Bhalerao, 1936 – облигатных паразитов дальневосточной черепахи [1].
Сведения о хозяевах и распространении трематод приведены по работам [7, 21, 28, 30, 34, 35, 38].
Систематический список трематод пресмыкающихся
Отряд Paramphistomida Skijabin et Schulz, 1937
Семейство Diplodiscidae Skrjabin, 1949
Кириллов Александр Александрович, к.б.н., с.н.с.; Кириллова Надежда Юрьевна, к.б.н., н.с., e-mail: [email protected]
Diplodiscus subclavatus (Pallas, 1760) Diesing, 1836
Хозяин: обыкновенный уж.
Локализация: кишечник.
Место обнаружения: Самарская область [10, 16].
Биология: Является случайным паразитом пресмыкающихся. Промежуточные хозяева – моллюски рода Planorbis. Окончательные хозяева – земноводные [21, 34].
Общее распространение: Европа.
Описание D. subclavatus от обыкновенного ужа (рис. 1): тело размерами 1,662-2,461X0,923-1,177 мм, имеет вид усеченного конуса, основанием которого служит большая брюшная (задняя) присоска. Ротовая присоска субтерминальная, 0,246-0,278X0,315-0,338 мм, на заднем конце имеет выступы – дивертикулы. Брюшная присоска большая, размерами 0,269-0,608X0,692-0,908 мм, располагается аборально. Пищевод длиной 0,154-0,308 мм, на заднем конце имеет небольшое вздутие (пищеводный бульбус) – 0,061-0,119X0,041-0,075 мм. Кишечные стволы широкие, простираются почти до уровня брюшной присоски.
Семенник один размерами 0,446-0,569X0,461-0,546 мм, лежит симметрично или слегка по диагонали в заднем конце тела. Половое отверстие располагается медианно позади бифуркации кишечника.
Яичник размерами 0,169-0,231 X 0,162-0,216 мм, лежит непосредственно позади семенника и сдвинут к одной из ветвей кишечника. Матка слабо извитая, образует петли впереди и позади яичника, у взрослых трематод заполнена яйцами размером 0,104-0,126X0,044-0,068 мм. Желточники овальной или неправильной формы фолликулами, расположены по бокам тела от уровня середины пищевода до брюшной присоски.
Отряд Plagiorchiida La Rue, 1957
Семейство Plagiorchiidae (Lühe, 1901) Ward, 1917
Plagiorchis elegans (Rudolphi, 1802)
Хозяин: прыткая и живородящая ящерицы, разноцветная ящурка, обыкновенный уж.
Локализация: кишечник.
Рис. 4. Neoglyphe sobolevi от прыткой ящерицы
Место обнаружения: Башкортостан [33], Чувашия [14], Нижегородская область [4, 5, 32]; Самарская область [9, 10, 14-16].Биология: Возможность паразитирования Р. elegans у холоднокровных животных, в частности у обыкновенного ужа ставилось под сомнения [31, 34].
Промежуточные хозяева – насекомые отрядов Díptera, Ephemeroptera, Odonata, ракообразные, моллюски рода Lymnaea [18, 19]. Для обыкновенного ужа является случайным хозяином. Широко распространенный паразит ящериц, птиц и грызунов [35].
Общее распространение: голарктический вид.
Описание Р. elegans от обыкновенного ужа (рис. 2): трематоды с удлиненно-овальным телом, длиной 1,611-1,788 мм, при ширине 0,438-0,495 мм. Большая часть тела покрыта шипиками, постепенно редеющими к заднему концу. Ротовая присоска субтерминальная, всегда больше брюшной, 0,172-0,196X0,163-0,181 мм. Брюшная присоска размерами 0,097-0,121X0,125-0,132 мм. Фаринкс округлый, 0,094-0,098 мм. Пищевод не выражен.
Рис.6. РгозоЮсш соп/шш от обык-новенного ужа
Кишечные стволы длинные, доходят до заднего конца тела.
Семенники округлые, 0,188-0,206X0,170-0,182 мм, залегают по диагонали в средней части тела. Задний семенник несколько больше переднего. Бурса цирруса удлиненная, 0,361-0,407 мм, расположена в зоне брюшной присоски, С-образно изогнута вдоль её боковой стороны. Семенной пузырек удлиненный, разделен перетяжкой на короткую переднюю и длинную заднюю части. Циррус длинный, невооруженный, размерами 0,125-0,136X0,131-0,144 мм.
Яичник округлый, диаметром 0,394-0,421 мм, больше брюшной присоски, расположен субмеди-ально между передним семенником и брюшной присоской. Желточники простираются от уровня фаринкса до заднего конца тела, где сливаются. В передней части тела желточные поля дорсально сливаются впереди брюшной присоски. Нисходящие и восходящие стволы матки залегают вен-трально, проходя между семенниками, образуют 8-образный изгиб. Яйца размерами 0,021-
Рис. 5. Pleurogenes claviger от обы-кновенного ужа
0,026X0,013-0,016 мм.
P. molini Lent et Freitas, 1940
Хозяин: прыткая и живородящая ящерицы.
Локализация: кишечник.
Место обнаружения: Самарская область [9, 10, 15].
Биология: Является специфичным паразитом ящериц. Жизненный цикл не изучен [35]. Можно предположить, что промежуточными хозяевами, как и других представителей рода Plagiorchis являются околоводные насекомые.
Общее распространение: палеарктический вид.
Astiotrema monticelli Stossich, 1904
Хозяин: обыкновенный и водяной ужи.
Локализация: кишечник.
Место обнаружения: Башкортостан [33], Мордовия (2008, личные данные), Татарстан [1, 23, 24], Самарская область [10].
Биология: Специфичный паразит ужеобразных змей. Промежуточные хозяева – моллюски Bithynia leachi, личинки и взрослые особи чесночницы, остромордой и озерной лягушки [35,36].
Общее распространение: Европа.
Leptophallus nigrovenosus (Bellingham, 1844) Lühe, 1909
Хозяин: обыкновенный уж, обыкновенная гадюка.
Локализация: пищевод, верхний отдел желудка.
Место обнаружения: Башкортостан [33], Мордовия (2008, личные данные) Татарстан [1], Самарская область [10], Ульяновская область [11].
Биология: Широко распространенный паразит ужеобразных и гадкжовых змей. Промежуточные хозяева – моллюски рода Lymnaea, амфибии и их личинки (остромордая, травяная, съедобная лягушки, жерлянка, жаба обыкновенная, тритон гребенчатый) [35].
Общее распространение: палеарктический вид.
Metaleptophallus gracillimus (Lühe, 1909) Yama-guti, 1958
Хозяин: обыкновенный уж.
Локализация: ротовая полость, пищевод.
Место обнаружения: Татарстан [1], Самарская область [10].
Биология: Обыкновенный и широко распространенный паразит ужеобразных и гадкжовых змей. Промежуточные хозяева – моллюск Planor-barius corneus, остромордая и травяная лягушки [35].
Общее распространение: Европа.
Opisthioglyphe ranae (Fröhlich, 1791)
Хозяин: обыкновенный уж.
Локализация: кишечник.
Место обнаружения: Мордовия (2008, личные данные), Татарстан [1, 23, 24], Самарская область
[10, 16].
Биология: Случайный паразит обыкновенного ужа. Отмечены как половозрелые, так и неполовозрелые особи паразита. Первыми промежуточными хозяевами являются моллюски рода Lymnaea, Planorbarius corneus, дополнительные – личинки комаров, веснянок, ручейников. Окончательные хозяева – земноводные. Метацеркарии могут встречаться у амфибий и их личинок. Встречается также у гадюк [34, 35].
Общее распространение: палеарктический вид.
Описание О. гапае от обыкновенного ужа (рис. 3): Тело удлиненно-овальное 1,162-1,385X0,465-0,585 мм, покрыто шипиками, исчезающими на уровне семенников. Ротовая присоска субтерминальная размерами 0,130-0,148X0,122-0,143 мм. Брюшная присоска 0,111-0,129X0,115-0,133 мм располагается преэкваториально. Фаринкс 0,063-0,074X0,069-0,077 мм. Пищевод 0,089-0,138 мм. Бифуркация кишечника на середине или около середины расстояния между присосками. Кишечные стволы длинные, относительно узкие, простираются к заднему концу тела, но до самого конца тела не доходят.
Семенники поперечно-овальные, иногда неправильной формы, цельнокрайние, располагаются один позади другого, тесно прилегая друг к другу. Передний семенник размером 0,111-0,247X0,119-0,148 мм, в большинстве случаев несколько меньше заднего – 0,133-0,248X0,148-0,207 мм. Бурса цирруса 0,128-0,172X0,070-0,087 мм, грушевидная или удлиненно-грушевидная, располагается между бифуркацией кишечника и брюшной присоской, под углом к продольной оси тела, может перекрываться присоской. Семенной пузырек извитый, занимает большую часть бурсы. Циррус короткий.
Яичник округлый или круглый размерами 0,130-0,155X0,122-0,148 мм, расположен справа у заднебокового края брюшной присоски. Желточ-ники занимают боковые поля тела, простираются от уровня пищевода или бифуркации кишечника до заднего конца тела, где, огибая слепые концы кишечных стволов, сливаются позади семенников. Матка относительно короткая. Петли матки занимают пространство между брюшной присоской и передним семенником. Половое отверстие субмедиальное, расположено на уровне бифуркации кишечника. Яйца крупные размерами 0,037-0,041X0,018-0,022 мм.
Paralepoderma cloacicola (Lühe, 1909)
Хозяин: обыкновенный уж, обыкновенная гадюка.
Локализация: прямая кишка.
Место обнаружения: Башкортостан [33], Мордовия (2008, личные данные), Татарстан [1, 23, 24], Самарская область [10, 11], Ульяновская область [11].
Биология: Один из наиболее обыкновенных и широко распространенных паразитов ужей и га-
дюк. Первыми промежуточными хозяевами являются моллюски Р1апогЫ$ р1апогЫя; вторые промежуточные хозяева – личинки и молодые особи озерной, остромордой, травяной лягушек и обыкновенной чесночницы [35].
Общее распространение: палеарктический вид.
Neoglyphe .\оЬо1е\ч (БсЬаЫуЫп, 1953) Уата§ий, 1958
Хозяин: прыткая ящерица.
Локализация: кишечник.
Место обнаружения: Самарская область [10, 16].
Биология: Это первая находка паразита у пресмыкающихся фауны России. Является случайным паразитом рептилий. Найдено всего два половозрелых экземпляра паразита. Жизненный цикл не изучен. Вероятные промежуточные хозяева – насекомые. Окончательными хозяевами служат представители семейства БойасЬе [35].
Общее распространение: голарктический вид.
Описание N. яоЬо1еУ1 от прыткой ящерицы (рис. 4): Трематоды грушевидной формы, с выемкой на заднем конце тела, размерами 0,535-0,576X0,301-0,312 мм. Тело покрыто шипиками, постепенно редеющими к заднему концу. Ротовая присоска субтерминальная, 0,045-0,048X0,049-0,052 мм. Брюшная присоска размерами 0,026-0,028X0,031-0,033 мм. Фаринкс 0,015-0,018X0,017-0,019 мм. Пищевод длиной 0,0150,021 мм. Бифуркация кишечника на середине расстояния между присосками. Кишечные стволы простираются к заднему концу тела, но самого конца не достигают.
Семенники неправильной формы, вытянуты поперек тела, располагаются постэкваториально, тесно прилегая друг к другу. Края семенников с выемками небольшой глубины. Передний семенник размерами 0,057-0,063X0,183-0,188 мм, задний – 0,083-0,087X0,172-0,177 мм. Бурса цирруса 0,240-0,251X0,034-0,037 мм, залегает под углом к продольной оси тела между передним краем переднего семенника и бифуркацией кишечника, С-образно изгибается проксимальным концом вокруг брюшной присоски.
Семенной пузырек большой, длина его более половины длины бурсы, разделен на короткую переднюю и удлиненную заднюю части. Циррус невооруженный длиной 0,108-0,113 мм. Половое отверстие субмедиальное.
Яичник размерами 0,098-0,105 X 0,078-0,084 мм, неправильной формы, с неглубокими выемками на краях, лежит субмедиально за брюшной присоской и половой бурсой. Желточники простираются по бокам тела от уровня бифуркации кишечника или чуть выше до заднего конца тела. В передней части тела желточники сливаются дорсально. Матка короткая, ее петли занимают пространство только между бурсой, передним семенником и яичником.
Дистальный конец матки с метратермом. Яйца 0,016-0,019X0,009-0,011 мм.
Семейство Macroderidae (Goodman, 1952) Ya-maguti, 1971
Macrodera longicollis (Abildgaard, 1788) Lühe, 1909
Хозяин: обыкновенный и водяной ужи.
Локализация: воздушный мешок легкого.
Место обнаружения: Мордовия (2008, личные данные), Татарстан [1], Самарская область [10].
Биология: Специфичный паразит ужеобразных змей. Является одним из самых обыкновенных и широко распространенных паразитов ужей. Первый промежуточный хозяин – моллюск Planorbis planorbis; второй промежуточный хозяин – амфибии (озерная и травяная лягушки) [30, 34].
Общее распространение: палеарктический вид.
Семейство Telorchiidae Looss, 1898
Telorchis assula (Dujardin, 1845)
Хозяин: обыкновенный и водяной ужи, обыкновенная гадюка.
Локализация: кишечник.
Место обнаружения: Башкортостан [33], Мордовия (2008, личные данные), Татарстан [1, 23, 24], Самарская область [10], Ульяновская область [11].
Биология: Специфичный паразит ужеобразных змей. Один из самых обычных и распространенных паразитов обыкновенного и водяного ужей, встречающийся практически в пределах всего ареала хозяев. Первый промежуточный хозяин -моллюск Planorbis planorbis. Вторыми промежуточными хозяевами являются головастики озерной и травяной лягушек. У гадюк и веретениц отмечался как случайный паразит [34, 35].
Общее распространение: палеарктический вид.
Семейство Encyclometridae Mehra, 1931
Encyclometra colubrimurorum Rudolphi, 1819
Хозяин: обыкновенный уж.
Локализация: нижний отдел пищевода, желудок.
Место обнаружения: Башкортостан [33], Татарстан [1, 23, 24], Самарская область [10].
Биология: Специфичный паразит ужеобразных змей. Один из типичных паразитов ужа обыкновенного, встречающийся на большей части ареала хозяина. Первый промежуточный хозяин неизвестен. Вторые промежуточные хозяева – земноводные: озерная, прудовая, остромордая лягушки, обыкновенная чесночница. У гадюк отмечается как случайный паразит [34, 35].
Общее распространение: палеарктический вид.
Семейство Pleurogenidae Looss, 1899
Pleurogenes с laviger (Rudolphi, 1819)
Хозяин: обыкновенный уж.
Локализация: кишечник.
Место обнаружения: Мордовия (2008, личные данные), Самарская область [10, 16].
Биология: Является случайным паразитом пресмыкающихся. Первые промежуточные моллюски – брюхоногие моллюски; вторые промежуточные хозяева – водные ракообразные, личинки и имаго околоводных насекомых. Окончательные хозяева -земноводные [34, 35].
Общее распространение: палеарктический вид.
Описание Р. claviger от обыкновенного ужа (рис. 5): Тело удлиненно-овальное размерами 1,631-3,185X0,602-0,997 мм, покрыто шипиками, которые постепенно редеют к заднему концу тела. Ротовая присоска субтерминальная, 0,200-0,268X0,222-0,305 мм. Брюшная присоска располагается преэкваториально размерами 0,146-0,188X0,153-0,191 мм. Фаринкс 0,088-0,109X0,101-0,138 мм. Бифуркация кишечника ближе к ротовой присоске. Кишечные стволы длинные и широкие, простираются постэкваториально, не достигают заднего конца тела. Заканчиваются на уровне переднего края семенников.
Семенники округлые или овальные, цельно-крайние размерами 0,196-0,247X0,212-0,267 мм, лежат симметрично или слегка по диагонали в заднем конце тела. Бурса цирруса 0,555-0,806X0,132-0,163 мм. Располагается под углом к продольной оси тела между брюшной присоской и левым краем тела. Семенной пузырек большой, извитый занимает около 1/3 длины бурсы. Семяиз-вергательный канал грушевидной формы, имеет толстые железистые стенки. Циррус короткий, 0,032-0,058 мм.
Яичник цельнокрайний, округлый, 0,1870,242 X 0,206-0,251 мм, располагается на одном или почти на одном уровне с брюшной присоской. Семяприемник лежит рядом с яичником непосредственно у заднебокового края брюшной присоски. Желточники располагаются в передней части тела, могут сливаться медиально на дорсальной стороне тела. Слева желточники простираются от уровня бурсы до нижнего края брюшной присоски или немного дальше. Справа желточники тянутся от уровня нижнего края ротовой присоски или бифуркации кишечника до уровня нижнего края брюшной присоски, могут переходить чуть ниже его. Матка образует многочисленные петли, которые располагаются в основном поперечно и занимают все пространство между брюшной присоской и задним концом тела.сиъ соп/шш (Ьоовв, 1894)
Хозяин: прыткая ящерица, обыкновенный уж.
Локализация: кишечник.
Место обнаружения: Татарстан [1], Самарская область [11, 15, 16].
Биология: Является случайным паразитом пресмыкающихся. Отмечены единичные находки неполовозрелых и половозрелых особей. Роль первого промежуточного хозяина выполняют брюхоногие моллюски. Вторые промежуточные хозяева -личинки и имаго околоводных насекомых. Окончательные хозяева – земноводные [34, 35].
Общее распространение: палеарктический вид.
Описание P. confusus от обыкновенного ужа (рис. 6): Тело округлое или овальное, размерами 0,786-1,037X0,553-0,814 мм, покрыто шипиками, постепенно редеющими к заднему концу. Ротовая присоска субтерминальная, 0,164-0,202X0,173-0,211 мм. Брюшная присоска размерами 0,142-0,151X0,129-0,135 мм, располагается постэкваториально. Фаринкс 0,046-0,064X0,053-0,071 мм. Пищевод короткий, длиной 0,033-0,054 мм. Бифуркация кишечника ближе к ротовой присоске. Кишечные стволы короткие и широкие не достигают уровня переднего края брюшной присоски.
Семенники овальные, размерами 0,164-0,185X0,133-0,148 мм, располагаются симметрично по обеим сторонам тела на уровне пищевода и бифуркации кишечника, снаружи от кишечных стволов. Бурса цирруса большая, булавовидная, 0,400-0,518X0,098-0,122 мм, расположена слева под небольшим углом к краю тела. Основание бурсы лежит на уровне брюшной присоски. В проксимальной части бурсы расположен крупный извитый семенной пузырек. Семяизвергательный канал грушевидный, занимает простатическую часть бурсы. Циррус невооруженный. Половое отверстие латеральное, на уровне пищевода, бифуркации кишечника или левого семенника.
Яичник размерами 0,129-0,147X0,151-0,192 мм, округлый, цельнокрайний расположен медиально или слегка субмедиально около переднего края брюшной присоски, между концами кишечных стволов. Желточники располагаются в передней части тела с обеих сторон ротовой присоски фа-ринкса, пищевода. Состоят из 15-18 крупных округлых фолликулов и медиально сливаются. Большая часть матки располагается позади брюшной присоски. Лишь небольшая ее часть извиваясь простирается в переднюю часть, располагаясь поперек тела вдоль нижнего края желточников. Дистальный отдел матки образует более или менее S-образно изогнутый метратерм. Яйца размерами 0,010-0,013X0,018-0,024 мм.
Отряд Strigeidida (La Rue, 1926) Sudarikov, 1959
Семейство Strigeidae Railliet, 1919
Strigea falconis Szidat, 1928, larvae
Хозяин: обыкновенный уж.
Локализация: брыжейка, жировая ткань.
Место обнаружения: Самарская область (2010, личные данные).
Биология: Обыкновенный уж является резерву-
арным хозяином. Первыми промежуточными хозяевами являются брюхоногие моллюски Planorbis planorbis и, в эксперименте, P. carinatus, Planor-barius corneus, Anisus vortex, A. leucostomus, A. con-tortus [37]. В роли вторых промежуточных хозяев выступают земноводные. Резервуарные хозяева -ужеобразные и гадюковые змеи. Взрослые гельминты паразитируют в кишечнике дневных хищных птиц отряда Falconiformis, реже в совиных [8, 21,26, 28, 34].
Общее распространение: космополит.
S. sphaerula (Rudolphi, 1803) Szidat, 1928, larvae
Хозяин: обыкновенный и водяной ужи, обыкновенная медянка.
Локализация: брыжейка, жировая ткань.
Место обнаружения: Башкортостан [33], Мордовия (2008, личные данные), Самарская область [10].
Биология: Впервые указывается для обыкновенной медянки. Является обычным паразитом змей (особенно ужей). В качестве первых промежуточных хозяев выступают брюхоногие моллюски; вторых – земноводные. Окончательные хозяева – врановые птицы [28, 30].
Рис. 7. Метацеркария Strigea sphaerula от обыкновенной медянки
Общее распространение: Европа.
Описание S. sphaerula, larvae от обыкновенной медянки (рис. 7): Метацеркарии заключены в цисту яйцевидной формы. Тело метацеркарии 0,4530,521 мм длиной, грушевидной формы, с сильно вздутым передним и относительно меньшим задним сегментом. Размеры переднего сегмента 0,365-0,408X0,325-0,358 мм, заднего – 0,088-0,112X0,169-0,263 мм. Вентральная впадина глубокая, карманообразная.
Ротовая присоска размерами 0,078-0,100X0,079-106 мм. Шаровидный фаринкс 0,041-0,044X0,042-0,047 мм. Пищевод короткий. Кишечные стволы слепо заканчиваются вблизи заднего конца тела. Латеральные псевдоприсоски хорошо развиты з длиной 0,174-0,203 мм, с крупными устьями, которые у фиксированных экземпляров округлые, широко открытые. Основания псевдоприсосок полушаровидной формы.
Брюшная присоска размерами 0,091-0,107X0,108-0,124 мм, лежит у дна вентральной впадины в пространстве между псевдоприсосками.
Орган Брандеса образован крупными лопастями -вентральной и дорзальной. Позади органа Брандеса располагается железистый комплекс в виде поперечно-овального тела размерами 0,056-0,071X0,114-0,146 мм.
S. strigis (Schrank, 1788) Abildgaard, 1790, larvae
Хозяин: обыкновенный и водяной ужи, обыкновенная и степная гадюки, прыткая ящерица.
Локализация: брыжейка, жировая ткань, полость тела.
Место обнаружения: Мордовия (2008, личные данные), Татарстан [1, 23, 24], Самарская область [10, 11], Ульяновская область [11].
Биология: Обычный и широко распространенный паразит змей фауны России. Значительная экстенсивность инвазии зафиксирована также у гадюки обыкновенной. У других рептилий метацеркарии встречаются редко. Первый промежуточный хозяин моллюск Planorbis planorbis. Вторые промежуточные хозяева – земноводные, которые могут быть и резервуарными хозяевами. Кроме них к резервуарным хозяевам относятся рептилии, птицы и млекопитающие. Окончательные хозяева – дневные хищные птицы и совы [28, 30].
Общее распространение: палеарктический вид.
Семейство Codonocephalidae (Sudarikov, 1959) Zhatkanbaeva, 1991
Codonocephalus urnigerus (Rudolphi, 1819) Diesing, 1850, larvae
Хозяин: обыкновенный уж.
Локализация: мышцы, полость тела, серозные покровы внутренних органов.
Место обнаружения: Татарстан [1, 23, 24].
Биология: Обыкновенный уж выступает в качестве резервуарного хозяина. Первыми промежуточными хозяевами являются брюхоногие моллюски рода Lymnaea. Земноводные служат вторыми промежуточными и резервуарными хозяевами. Мариты паразитируют в кишечнике большой и малой выпей, серой цапли. Резервуарными хозяевами кроме ужеобразных являются гадюковые змеи [21,26, 27, 34].
Общее распространение: палеарктический вид.
Семейство Alariidae (Hall et Wigdor, 1918) Tubangui, 1922
Alaria alata (Goeze, 1782), larvae
Хозяин: обыкновенный уж, обыкновенная и степная гадюки, обыкновенная медянка.
Локализация: жировая ткань, полость тела.
Место обнаружения: Башкортостан [33], Мордовия (2008, личные данные), Татарстан [1, 23, 24], Самарская область [10, 11], Ульяновская область [и].
Биология: Один из самых распространенных и многочисленных паразитов змей. Чаще всего А. alata встречается у ужей. Первыми промежуточ-
ными хозяевами являются брюхоногие моллюски. Роль резервуарных хозяев выполняют амфибии, рептилии, дневные хищные, совиные и куриные птицы, микромаммалии. Окончательные хозяева -представители семейств псовых (лисица, енотовидная и домашняя собаки и др.) и куньих (американская норка) [20, 22, 28].
Общее распространение: космополит.
Pharyngostomum cor datum (Diesing, 1850) Ciurea, 1922, larvae
Хозяин: обыкновенный и водяной ужи.
Локализация: жировая ткань, серозные покровы внутренних органов.
Место обнаружения: Мордовия (2008, личные данные), Самарская область [10].
Биология: Обычные и широко распространенные паразиты змей фауны России. Ужи являются резервуарными хозяевами. Первый промежуточный хозяин – моллюск Planorhis planorhis. Вторыми промежуточными хозяевами являются земноводные. К резервуарным хозяевам, кроме рептилий, относятся врановые птицы, насекомоядные и грызуны. Окончательные хозяева – хищные млекопитающие семейств кошачьих и псовых, которые могут выступать и в роли резервуарных [29, 34].
Общее распространение: палеарктический вид.
Семейство Diplostomidae Poirier, 1886
Neodiplostomum spathoides Dubois, 1937, larvae
Хозяин: обыкновенный и водяной ужи.
Локализация: жировая ткань, серозные покровы внутренних органов.
Место обнаружения: Самарская область (2010, личные данные).
Биология: Ужи выступают в качестве резервуарных хозяев. Первыми промежуточными хозяевами являются брюхоногие моллюски рода Planorbis. Земноводные служат вторыми промежуточными хозяевами. Мариты паразитируют в кишечнике дневных хищных птиц отряда Falconifor-mes. Резервуарными хозяевами служат ужеобраз-ные и гадюковые змеи [21, 28, 34].
Общее распространение: палеарктический вид.
Анализ трематодофауны пресмыкающихся Среднего Поволжья показал, что большинство имагинальных форм паразитов представлено специфичными видами. К узко специфичным относятся у обыкновенного и водяного ужей: Е. colu-hrimurorum,M. longicollis, P. cloacicola, T. assula; y прыткой и живородящей ящериц – Р. тоlini. Специфичными паразитами ужеобразных и гадкжовых змей являются A. monticelli, L. nigrovenosus, M. gracillimus.
P. elegans паразитирует y представителей разных классов позвоночных животных: птиц, млекопитающих (летучие мыши, грызуны) и рептилий (ящериц) [6, 34, 35]. Обычный паразит ящериц. У
обыкновенного ужа встречается случайно. По всей видимости, заражение змеи этим паразитом произошло при потреблении (случайном вместе с пищей, либо специальном, как пищевого объекта) промежуточного хозяина гельминта – личинок и имаго стрекоз, ручейников.
К случайным паразитам рептилий следует отнести I). 5иЪс1ауа1ш, Р. claviger, Р. соп/шш, О. гапае. Эти специфичные паразиты амфибий попадают в организм обыкновенного ужа, безусловно, при питании последнего бесхвостыми земноводными. В условиях Среднего Поволжья у обыкновенного ужа амфибии составляют более 80% рациона, у водяного ужа – 10% [2]. Но, вероятно, заражение змей происходит и при заглатывании (случайном или целенаправленном) вторых промежуточных хозяев этих гельминтов – личинок амфибий, моллюсков семейства Lymnaeidae, циклопов, личинок и имаго околоводных насекомых. На Самарской Луке отмечены случаи попыток водяного ужа заглотить обыкновенного прудовика Ьутпаеа stagnalis (личные наблюдения). Вопрос требует дальнейшего изучения. Заражение прыткой ящерицы Р. соп/ияия, конечно, произошло в околоводных стациях вследствие питания рептилии личинками и имаго околоводных насекомых.
N. яоЬо1еу1 отмечается в качестве случайного паразита прыткой ящерицы. Найдено всего 2 зрелых экземпляра паразита [10, 16]. Окончательными хозяевами трематоды являются обыкновенная и малая бурозубки. Заражение рептилии произошло при потреблении вероятных промежуточных хозяев гельминта – околоводных насекомых [35].
Личиночными формами трематод 5″. falconis. Б. $р1гаеги1а, Б. strigis С. urnigerus, А. аШа, Р1г. corda-Шт и N. spathoides рептилии инвазируются двумя путями трофическим и топическим, по классификации В.Л. Контримавичуса [17]. Основной путь, по-видимому, топический, когда тесная связь змей (обыкновенного и водяного ужей и, отчасти, обыкновенной гадюки) с водной средой обусловливает заражение их церкариями трематод, выходящих из моллюсков. Проникновение их в рептилий происходит через слизистую ротовой полости и клоаку, поскольку покров тела пресмыкающихся препятствует [30]. Второй путь – трофический, когда при питании змей бесхвостыми амфибиями еще не закончившие своего развития (неинцисти-рованные) метацеркарии из организма проглоченных амфибий проходят через стенку кишечника пресмыкающихся и локализуются на своем обычном месте. Уже инцистированные метацеркарии не имеют перфораторных органов и желез проникновения и неспособны «переходить» из амфибий в змей [30].
Наиболее разнообразна фауна трематод у обыкновенного ужа (19 видов), что является следствием его околоводного образа жизни и питания бесхвостыми амфибиями, являющимися вторыми промежуточными хозяевами большинства видов
трематод змеи.
Видовой состав трематод водяного ужа и обыкновенной гадюки (7 и 5 видов, соответственно) представляет собой сильно обедненную тремато-дофауну обыкновенного ужа. Все обнаруженные у водяного ужа и обыкновенной гадюки виды трематод встречаются и у обыкновенного ужа, что связано со спецификой питания и стационным размещением этих рептилий. Водяной уж является преимущественно ихтиофагом (рыба составляет 90% рациона) и большую часть времени проводит в воде на удалении от прибрежной зоны. Обыкновенная гадюка предпочитает разнообразные стации, где обитают основные пищевые объекты змеи – мышевидные грызуны (60-100% рациона в разных популяциях). Рептилия меньше времени проводит в водной среде, предпочитая, находится на берегу [2].
Трематодофауна степной гадюки и обыкновенной медянки представлена исключительно личиночными формами (по 2 вида) и объясняется, с одной стороны, обитанием в сухих стациях и своеобразием пищевого рациона; с другой – малым числом исследованных животных – по 7 особей [10,11].
Среди ящериц наибольшее число видов трематод отмечается у прыткой ящерицы (5 видов), что связано с многообразием стаций обитания рептилии и изменением спектра питания ящерицы в разных местообитаниях, поскольку состав пищи животного отражает численное соотношение насекомых и других беспозвоночных в каждой конкретной стации.
Бедна в качественном отношении трематодофауна живородящей ящерицы, у которой зарегистрировано всего 2 вида, Это связано, с одной стороны, с меньшими размерами тела и челюстей живородки, которые уменьшают спектр ее питания, исключая крупную добычу; с другой – с малым числом исследованных животных [9, 10].
Лишь один вид трематод P. elegans отмечен у разноцветной ящурки. Что, с одной стороны, связано с малым числом исследованных ящериц (всего 15 особей), с другой, тем, что исследованная популяция ящериц находится на северной границе ареала (Кириллов, Епланова, 2005 а, б).
Не были обнаружены трематоды у ломкой веретеницы [10]. Таким образом, по нашим и литературным данным у пресмыкающихся Среднего Поволжья зарегистрирован 21 вид трематод. Всего для рептилий фауны России к настоящему времени известно 39 видов трематод [9, 10, 11, 33, 34].
Нами впервые для пресмыкающихся фауны России указывается трематода Neoglyphe sobolevi (хозяин – прыткая ящерица). Для Strigea sphaerula, larvae отмечен новый хозяин – обыкновенная медянка. Разные виды рептилий указываются в качестве случайных хозяев для 6 видов гельминтов: Diplodiscus subclavatus, Plagiorchis elegans, Pleuro-
genes claviger, Prosotocus confusus, Opisthioglyphe ranae, Neoglyphe sobolevi.
Дальнейшие перспективы гельминтологического изучения пресмыкающихся Среднего Поволжья связаны в первую очередь, с расширением районов исследований, поскольку на карте региона существует еще много «белых пятен» в отношении иссле-дованности паразитов рептилий. Лимитирующий фактор исследований в данном направлении – создание в регионах Среднего Поволжья Красных книг, куда включаются многие виды пресмыкающихся.
СПИСОК ЛИТЕРАТУРЫ
1. Аль-Завахра Х.А. Змеи Татарстана: Автореф. дис. … канд. биол. наук. Казань, 1992. 18 с.
2. Бакиев А.Г., Гаранин В.И., Литвинов H.A., Павлов A.B., Ратников В.Ю. Змеи Волжско-Камского края. Самара: Изд-во Самарского НЦ РАН, 2004. 192 с.
3. Бакиев А.Г., Маленев A.JI. Пресмыкающиеся Среднего Поволжья. Тольятти: ИЭВБ РАН, 1996. 25 с.
4. Борисова В.И. Основные закономерности распределения паразитов ящериц рода Lacerta L. // Наземные и водные экосистемы. 1981. Вып. 4. С.115-120.
5. Борисова В.И., Фадеева Г.А. Эколого-паразитологические исследования прыткой ящерицы в различных экосистемах // Наземные и водные экосистемы. 1990. Вып. 13. С.3441.
6. Демидова Т.Н., Вехник В.П. Трематоды (Trematoda, Monorchiidae) ночниц Myotis brandtii и М. mystacinus (Chiroptera, Vespertilionidae) Самарской Луки (Россия) // Вестник зоол. 2004. Т. 38. Вып. 5. С. 71-74.
7. Иванов В.М., Семенова H.H. Видовой состав и экологические особенности трематод рептилий дельты Волги // Паразитология. 2000. Т. 34. Вып. 3. С. 228-233.
8. Пскова Н.И., Корнюшин В.В., Буканева Н.С. К изучению плоских червей хищных птиц (Falconiformes) и сов (Strigiformes) Украины // IX конф. Укр. паразитол. общества: Тез. докл. Ч. 2. Киев: Наукова думка, 1980. С. 96-98.
9. Кириллов A.A. Гельминтофауна настоящих ящериц (Lacertidae, Lacerta) Бузулукского бора и Красносамар-ского лесничества // II конф. герпетологов Поволжья: Тез. докл. Тольятти: ИЭВБ РАН, 1999. С. 26-27.
10. Кириллов A.A. Фауна гельминтов пресмыкающихся Самарской области // Изв. Самарского НЦ РАН. 2000. № 3. С. 324-329.
11. Кириллов A.A., Бакиев А.Г. К изучению гельминтофау-ны гадюковых (Viperidae) Среднего Поволжья // Самарская Лука. Бюлл. 2003. № 13. С. 331-336.
12. Кириллов A.A., Евланов H.A. Особенности формирования гельминтофауны обыкновенного ужа Natrix natrix в зависимости от размерной структуры // Актуальные проблемы герпетологии и токсинологии: Сб. науч. тр. Вып. 3. Тольятти: ИЭВБ РАН, 1999 а. С. 73-76.
13. Кириллов A.A., Евланов H.A. Характеристика гельминтофауны обыкновенного и водяного ужей Самарской Луки // Самарская Лука на пороге третьего тысячелетия: мат-лы к докл. «Состояние природного и культурного наследия Самарской Луки». Тольятти: ИЭВБ РАН-ОСНП Парквей, 1999 б. С. 204-205.
14. Кириллов A.A., Епланова Г.В. Рельминтофауна синтопи-ческих популяций ящериц Среднего Поволжья // Актуальные проблемы герпетологии и токсинологии. Вып. 8. Тольятти: ИЭВБ РАН, 2005 а. С. 57-59.
15. Кириллов A.A., Епланова Г.В. Гельминтофауна ящериц (Sauria) Самарской области // Актуальные проблемы
герпетологии и токсинологии. Вып. 8. Тольятти: ИЭВБ РАН, 2005 б. С. 60-66.
16. Кириллов A.A., Кириллова Н.Ю. Паразитирование у рептилий Самарской области гельминтов, свойственных другим животным // Актуальные проблемы герпетологии и токсинологии. Вып. 10. Тольятти: ИЭВБ РАН , 2007. С.70-72.
17. Контримавичус В.Л. Гельминтофауна куньих и пути её формирования. М.: Наука, 1969. 430 с.
18. Краснолобова Т.А. Принципы систематики трематод рода Plagiorchis Luhe, 1899 // Цестоды и трематоды: Тр. ГЕЛАН. 1977. Т. 27. С. 65-110.
19. Краснолобова Т.А. Изучение биологических особенностей Plagiorchis elegans в дельте Волги // Тр. ГЕЛАН СССР. 1979. Т. 29. С. 75-80.
20. Потехина Л.Ф. Цикл развития возбудителя аляриоза лисиц и собак // Тр. Всесоюз. ин-та гельминтологии. 1950. Вып. 4. С. 7-17.
21 .Рыжиков K.M., Шарпило В.П., Шевченко H.H. Гельминты амфибий фауны СССР. М.: Наука, 1980. 279 с.
22. Савинов В.А. Особенности развития Alaria alata (Goeze, 1782) в организме дефинитивного и резервуарного хозяев // Работы по гельминтологии. М.: Изд-во АН СССР, 1953. С. 611-616
23. Смирнова М.И. Биоценотические связи гельминтов некоторых позвоночных животных побережья Куйбышевского водохранилища: Автореф. дисс. … канд. биол. наук. Казань, 1970. 17 с.
24. Смирнова М.И. Гельминтофауна обыкновенного ужа Саратовского участка Волжско-Камского заповедника // Природные ресурсы Волжско-Камского края. Животный мир. 1971. Вып. 3. С. 164-167.
25. Смирнова М.И., Алъ-Завахра Х.А. Новые данные о гель-минтофауне ужа обыкновенного в Татарской ССР. Деп. В ОНП НПЭЦ «Верас-эко» и ИЗ АН Беларуси. 21.08.1991. №91-14.
26. Смогоржевская Л.А. Гельминты водоплавающих и болотных птиц фауны Украины. Киев: Наукова думка, 1976.416 с.
27. Судариков В.Е. Отряд Strigeidida (La Rue, 1926) Su-darikov, 1959 II Скрябин К.И. Трематоды животных и
человека. Т. 16. Ч. 1. М.: Изд-во АН СССР, 1959. С. 219-631.
28. Судариков В.Е. Отряд Strigeidida (La Rue, 1926) Su-darikov, 1959 II Скрябин К.И. Трематоды животных и человека. Т. 24. Ч. 5. М.: Наука, 1971. С. 69-272.
29. Судариков В.Е., Ломакин В.В., Семенова H.H. Трематода Pharyngostomum cordatum (Alariidae, Hall et Wigdor, 1918) и её жизненный цикл в условиях дельты Волги // Гельминты животных. М.: Наука, 1991. С. 142-147.
30. Судариков В.Е., Шигин A.A., Курочшн Ю.В., Ломакин В.В., Стенько Р.П., Юрлова НИ. Метацеркарии трематод – паразиты пресноводных гидробионтов Центральной России. М.: Наука, 2002. 298 с.
31. Тимофеева Т.Н. Диагностическое значение некоторых признаков видов рода Plagiorchis Luhe, 1899 II Гельминты человека, животных, растений и борьба с ними. М. : Изд-во АН СССР, 1963.С. 99-106.
32. Фадеева Г.А. Экологические аспекты формирования и стабильности паразитарных систем на примере прыткой ящерицы: Автореф. дисс. … канд. биол. наук. Н. Новгород, 2000. 21 с.
33. Хабибуллин В.Ф. Пресмыкающиеся республики Башкортостан: Автореф. дисс. … канд. биол. наук. Уфа, 1999. 18 с.
34. Шарпило В.П. Паразитические черви пресмыкающихся фауны СССР. Киев: Наукова Думка, 1976. 376 с.
35. Шарпило В.П., Искова Н.И. Фауна Украины. Трематоды. Плагиорхиаты (Plagiorchiata). 34 (3). Киев: Наукова Думка, 1989.280 с.
36. Шевченко H.H., Вергун Г.И. Расшифровка цикла развития трематоды Astiotrema monticelli Stossich 1904 II Докл. АН СССР. 1960. Т. 130. №4. С. 949-952.
37. Odening К. Die lebenszyklen von Strigea falconispalumbi (Viborg), S. strigis (Schrank) und S. sphaerula (Rudolphi) (Trematoda, Strigeida) im Raum Berlin II Zool. Yahrb. Syst. 1967. H. 94. S. 1-67.
38.Prudhoe S., Bray R.A. Platyhelminth parasites of the Amphibia. London: British Museum (Natural History) and Oxford Univ. Press, 1982. 217 p.
TREMATODES (TREMATODA) OF REPTILES FROM MIDDLE VOLGA REGION
© 2011 A.A. Kirillov, N.Yu. Kirillova
Institute of Ecology of Volga river Basin ofRAE, Togliatti
Data on all known currently trematodes, revealed in reptiles of the Middle Volga are generalised and analysed in article. 21 species of trematodes is marked from reptiles of the Middle Volga region. We underline trematode Neogly-phe sobolevi for the first time for reptiles of fauna of Russia. The new host – smooth snake is noted for larva’ Strigea sphaerula. Different species of reptiles are specified as casual hosts for 6 trematodes. Descriptions and original drawings of these parasites are brought. Key words: trematodes, reptiles, Middle Volga region.
Kirillov Alexander Aleksandrovich, Candidate of Biology, senior researcher; Kirillova Nadezhda Jurevna, Candidate of Biology, senior researcher e-mail: [email protected]
Трематоды
Трематоды (Trematoda) представлены примерно 6000 видов, все из которых являются паразитами животных и человека. Для трематод характерно непрямое развитие, причем их жизненные циклы довольно сложны и связаны со сменой хозяев. По этой причине их называют биогельминтами. Циклы развития связаны с циркуляцией в организме постоянных хозяев (дефинитивных, окончательных) мариты (взрослой формы) и в организме промежуточных хозяев личиночных форм (рис. 23). Дефинитивными хозяевами трематод являются человек и другие животные.[ …]
Первым отечественным исследователем личинок трематод – метацеркарий, развивающихся в организме стрекоз, был Д.Ф. Синицын /1905/. Опыты по выяснению жизненных циклов трематод у рыб и лягушек он проводил в окрестностях Варшавы. Скармливая церкарии нескольких видов трематод личинкам водных насекомых, главным образом стрекоз, он выяснил, что дополнительными (или вторыми промежуточными) хозяевами трематод рода G-orgode— га являются личинки стрекоз рода Epitheca, хозяевами трематод Sicrjabinoeces similis (автор ошибочно привел ее под названием Haematoloechus variegatus ) и Halipegus ovo — caudatus — личинки и имаго стрекоз Calopteryx virgo; мета— перкарии трематоды Prosotocus confusus Д.Ф. Синицын находил в личинках и взрослых Aeschna sp.; Pleurogenoides medians — в жировом теле и полости тела Coenagrion sp. Для всех обнаруженных метацеркарий трематод автор дал морфологическое описание и рисунки.[ …]
У всех трематод в первом промежуточном хозяине — моллюске — формируются (как у всех трематод) церкарии, которые выходят в воду, внедряются в подкожную клетчатку и мышцы рыб. У метагонимусов и эхинохазмусов личинки поражают жабры, плавники и чешую рыб, у меторхисов — жабры и мускулатуру рыб. В местах локализации личинок образуются метацеркарии, которые по истечении 35 сут становятся инвазионными. У последней трематоды диаметр метацеркариев равен примерно 0,18—0,21 мм.[ …]
Марита трематоды имеет продолговатое тело, заметно суживающееся к переднему концу, длиной 8—13 мм и шириной 1,5—2,5 мм.[ …]
Взрослые трематоды, находясь в местах локализации — желчных ходах и реже в поджелудочной железе окончательных хозяев, продуцируют яйца, которые с фекалиями выделяются наружу. Во внешней среде яйца рассеиваются и заглатываются моллюсками. В их кишечнике из яйца вылупляется мирацидий, мигрирующий в печень и превращающийся в спороцисту. Срок развития от яйца до мета-церкария составляет 2,5 мес. Из двенадцатиперстной кишки дефинитивных хозяев паразиты проникают через протоки в печень и поджелудочную железу. Описторхисы в организме дефинитивного хозяина достигают половой зрелости через 21—28 сут. Продолжительность жизни гельминта в плотоядных животных более 3 лет, в организме человека — 10—20 лет.[ …]
Метацеркарий трематоды округло-овальной формы, длиной 0,23—0,63 мм и шириной 0,12—0,28 мм, серого цвета. В свежеприготовленных препаратах у метацеркариев хорошо заметны ротовая и брюшная присоски, в задней части экскреторный пузырек округлой формы, заполненный гранулами черного цвета (рис. 68).[ …]
Это продолговатые трематоды, у которых бока слегка втянуты внутрь (талии). Поверхность гельминта покрыта шипами. Брюшная присоска отсутствует, но на уровне середины тела с правой стороны находится половая присоска. Диаметр ротовой присоски 0,015—0,017 мм. Два круглых семенника 10 расположены в задней части тела наискосок один к другому. Овальный яичник находится впереди семенников. Матка, заполненная яйцами, занимает всю заднюю часть паразита. Желточники 7 в виде глыбок локализованы сбоку в задней части паразита (см. рис. 70, Г).[ …]
Гинецинская Т. А. Трематоды, их жизненные циклы, биология и эволюция. Л.: Наука. 1968.411 с.[ …]
Это очень мелкие трематоды, которым свойствен прогенез на стадии метацеркарии и быстрое созревание в окончательном хозяине. Продолжительность их пребывания в рыбе исчисляется, по-видимому, днями или неделями. Благодаря этому трематоды исчезают у рыб вскоре после того, как рыбы перестают питаться, что происходит в конце осени. Возможно, такая стратегия позволяет избегать конкуренции с более крупными и долго созревающими видами, поскольку летом основной предполагаемый конкурент (S. globiporum) имеет мелкие размеры и еще низкую численность. Развитие нематоды С. tomentosa может происходить как с участием олигохет, так и вообще без промежуточного хозяина (Ломакин, Тро-фименко, 1982). Половой зрелости в рыбе личинки достигают быстро – через 2 мес. Принадлежность С. tomentosa к “летним” видам можно объяснить той же причиной, что и для Р. тагке-witschi.[ …]
Анатомия и биология трематод. Это плоские черви листовидной формы. Тело обычно удлиненно-овальное, размером от 1 мм до 15 см. Оно покрыто тегументом (гладкое, с шипами или с щетинками), состоящим из наружного и внутреннего слоев (рис. 61). У большинства трематод две присоски: ротовая (на переднем конце тела) и брюшная (чаще на середине тела). У некоторых трематод, например у представителей Бап шшсоШае, паразитирующих в крови рыб, присоски рудиментированы или отсутствуют. От ротового отверстия отходит глотка, затем пищевод, переходящий в кишечник. Последний в виде двух стволов, достигнув заднего конца тела, слепо заканчивается. В очень редких случаях кишечные стволы открываются наружу. Сосальщики питаются тканевой жидкостью, слизью, некоторые — кровью. Продукты обмена выделяются через экскреторную систему протонефридиального типа. По всему телу размещены канальцы, объединяющиеся в два выделительных канала, открывающихся на заднем конце тела.[ …]
У цестод в отличие от трематод (в стадии имаго) наружный слой гегумента снабжен микроворсинками, играющими большую роль в питании цестод (см. рис. 61). Как правило, на дистальных и проксимальных концах ворсинок и на цитоплазматической мембране цестод адсорбируются пищеварительные ферменты хозяина, способные расщеплять белки, углеводы и жиры до конечных мономеров, которые в последующем транспортируются в организм паразита.[ …]
В местах локализации трематоды продуцируют яйца, которые при выходе из матки попадают в кровеносное русло. Затем с током крови они разносятся по всему телу и задерживаются в мельчайших капиллярах различных органов, в том числе в жаберном аппарате и в почках. Из яиц, застрявших в капиллярах жабр, выходят мераци-дии, при помощи стилета в передней части тела разрывают капилляры и попадают в воду. В воде они активно плавают в поисках промежуточных хозяев. Спороциста (мешкообразное образование) и редий с удлиненным телом, имеющим ротовое отверстие и короткий мешковидный кишечник, характерны для многих личинок трематод. Церкарии формируются в редиях и в конечной стадии развития имеют удлиненное тело с вилообразно раздвоенным в конце хвостом и высоким прозрачным гребнем на дорсальной стороне передней части тела. Церкарии покидают тело моллюска через мантийную полость и в воде (в теплый сезон года) активно отыскивают рыб—дефинитивных хозяев, внедряются в жабры и кожный покров, попадая в кровеносные сосуды, где достигают половой зрелости (рис. 66). Сроки развития личиночных стадий в моллюске до церкария и трематод до половозрелости у рыб почти не изучены.[ …]
Характерным признаком трематоды является тупо срезанный задний конец, который втянут внутрь в виде большой присоски. Два округлой формы семенника 6 расположены в заднем конце тела и как бы охватывают два толстых ствола кишечника, заканчивающихся тупо. Матка 4 занимает среднюю часть тела; на ее уровне по бокам тела вдоль кишечных стволов находятся желточники (см. рис. 70, Б).[ …]
Размашкин Д. А. О личинках трематод, паразитирующих у рыб водоемов Обь-Иртышского бассейна // Болезни и паразиты рыб Ледовитоморской провинции (в пределах СССР).[ …]
Илюшина ТЛ. Т. 29. -С 59-66.[ …]
Краснолобова Т.А. Биологический цикл трематоды Prosthogonimus pel— lucidus (Lunstow, 1B73), возбудителя заболевания птиц // Helminthologie. – М.: Изд-во АН СССР. – 1962. – Т. 4. – С. 217-229.[ …]
Систематика и краткая характеристика трематод. Трематодозы — инвазионные заболевания, возбудителями которых являются различные виды трематод или их личинки, относящиеся к классу трематод — Тгеташёа, или дигенетических сосальщиков, типу РМЬ гтпШез.[ …]
Шевченко H.H., Вергун Г.И. О жизненном цикле трематоды амфибий Pro— sotocus confusus (Looss, 1894) Looss, 1899 // Helmintholo— gica. – 1961. – T. 3, вып. 1-4. – C. 294-298.[ …]
Новосибирск: Наука.[ …]
Из чисяа инвазированных особей метацеркариями одного вида трематод S. sanguineum составляли 68,55, a S. vulgatum- 70,02 %, двумя видами – соответственно 23, 38 и 23,34 %, тремя – 8,46 и 6,36 %, четырьмя – 1,61 и 1,06 % и пятью – 0,40 и 0,00 %, т.е. эти соотношения для обоих видов стрекоз очень близки. Если принять всех инвазированных стрекоз за 100 %, то инвазированные одним видом трематод составят около 70 %, двумя – примерно в 3 раза меньше, тремя – также примерно в 3 раза меньше, чем двумя, и четырьмя – в 5-6 раз меньше, чем тремя. Эти соотношения позволили выяснить некоторые формы взаимоотношений паразитов со стрекозами-хо— зяевами /Павлюк, 19836/.[ …]
Илюшина ТЛ.[ …]
С.В. Карпенко и В.В. Заика /1979/ в Северной Кулунде изучали экологию трематоды Plagiorchis elegans. Наибольшая экстенсивность инвазии метацеркариями отмечена у стрекоз S. vulga— tum – 46,7 и S. flaveolum – 44 %, т.е. точно такая же, как и по данным Т.Л, Илюшиной /1973/ для Карасукских озер.[ …]
У личинок Aeschna sp. из дельты Дуная Т.Н. Комарова /1968/ выявила метацеркарии трематоды Pleurogenoides medians.[ …]
В.Н. Крылова /1969/ для этой же территории в качестве дополнительных хозяев трематод рода Prosthogonimus отмечает Les— tes dryas, Sympetrum flaveolum, S. scoticum и E. cy— athigerum, т.е. те же виды, в которых вышеуказанные авторы находили плягиорхид. ЭИ личинок стрекоз в разных водоемах составляла от 10 до 70 %. Автор сообщает, что максимумы интенсивности и экстенсивности инвазии приходятся на июнь и июль, т.е. на время личиночного развития стрекоз.[ …]
В Ковровском районе Владимирской области Т.А. Краснолобова /1958, 1962/ проводила исследования биологии трематоды Prost— hogonimus pellucidus (= P. cuneatus), возбудителя заболевания яйцеводов домашних птиц – простогонимоза. Из 11 исследованных видов стрекоз метал ер кари ями этой трематоды были заражены: Cor— dulia aenea, Libellula quadrimaculata, Leucorrhinia rubicunda, Somatochlora metalliaca. Выявленные у стрекоз метацеркарии были представлены тремя типами: в двухслойной, однослойной цисте и свободные (неинцистированные). Автор дала описание всех трех типов метацеркарий. В дальнейшем Т.А. Краснолобовой /1968/ установлено, что на территории СССР существуют два вида из рода Prosthogonimus — Р. cuneatus и P. ovatus, а все другие названия являются лишь их синонимами.[ …]
На территории Астраханского заповедника Т.А. Краснолобова /1979/, проводившая исследования цикла развития трематоды Р1а— giorchis el gans, выяснила, что найденные у E. cyathigerum метацеркарии полностью идентичны описанным В.Е. Судариковым и Е.М. Кармановой от слепня под названием Р. laricola.[ …]
Выше города по течению реки у рыб обнаружено 16 видов паразитов. Наиболее часто встречаются мета-церкарии трематод Diplostomum spathaceum, Opisthorchis felineus, Paracoenogonimus ovatus, Prochemistomum hughesi, Prochemistomum sp. и ракообразные Tracheliastes polycolpus. У рыб из p. Туры, отловленных в черте Тюмени, количество видов паразитов то же, но видовой состав отличается отсутствием ракообразных, цестод Triaenophorus nodulosus, Caryophyllaeus laticeps, ряда трематод и наличием цестод Caryophyllaeides fennica, трематод Cotylurus variegates, Allocreadium isoporum и нематод Raphidascaris acus. Ниже города по течению реки у рыб найдены только трематоды – 14 видов. Различия видового состава паразитов рыб в разных участках реки объясняются особенностями гидрологического режима и состава гидроценоза в каждом конкретном пункте. Выпадение из паразитофауны эктопаразитов Т. polycolpus и цестод Т. nodulosus, жизненный цикл которых связан с веслоногими раками, при протекании реки по городу может свидетельствовать о наличии в воде вредных примесей, к которым эти группы чувствительны.[ …]
Простейших окрашивают по Романовскому — Гимзе, метиленовым синим или железным гематоксилином. Для окраски трематод и цестод применяют квасцовый кармин. Моногенетических сосальщиков, миксоспоридий заключают в глицерин-желатину, замазывают сухие края покровных стекол бальзамом или черным лаком и хранят. Скребней и рачков также не красят, а для приготовления постоянных препаратов просветляют и заливают в бальзам. Окраску и заключение паразитов рыб проводят по общепринятым в паразитологии методикам (см. специальные руководства).[ …]
Голикова МЛ.[ …]
Так, в прудах разводят ветвистоусых рачков, элиминирующих свободноживущие стадии паразитов. Используют для выращивания в поликультуре рыб моллюскофагов, например черного амура, сигов, бентофагов, карпа старших возрастных групп, уничтожающих моллюсков — промежуточных хозяев некоторых гельминтов. Вселяют белого амура, питающегося высшими водными растениями и ухудшающего экологические условия обитания и размножения моллюсков — промежуточных хозяев возбудителей трематодозов. В борьбе с цестодозами, например лигулезом, в озерные хозяйства можно вселять невосприимчивых рыб, например сиговых. При инфекционных заболеваниях также применяют совместное выращивание рыб, восприимчивых и невосприимчивых к данному заболеванию. Так, в хозяйствах, неблагополучных по аэромонозу, в поликультуре с карпом содержат растительноядных рыб. Биологические методы борьбы являются составной частью экологически обоснованной профилактики заболеваний рыб, которая основана на управлении экологическими факторами и процессами в рыбохозяйственном водоеме.[ …]
При гельминтологическом обследовании рыб в мышцах, в других органах и тканях нередко обнаруживают личинок либо зрелых трематод. При этом отдельные из них в случае высокой интенсивности заражения могут приводить к значительной потере или снижению продуктивности рыб, ухудшению качества мяса и икры. Например, в кишечнике рыб таких видов, как плотва, карась, щипов-ка, лещ, и некоторых других карповых обнаруживают трематоды Allocreadium isoporum, относящиеся к семейству Allocreadiidae. Длина гельминта 0,9—3,6 мм, ширина 0,3—0,8 мм. Он сероватожелтоватого или розоватого цвета. Снабжен двумя присосками, характерными для трематод. У него два семенника, расположенные один за другим в задней части тела. Желточники находятся по бокам задней половины тела. Развитие происходит с участием пресноводных моллюсков рода Sphaerium — промежуточных хозяев. Дополнительными хозяевами являются личинки насекомых — Ephemera, Anobolia, Chaetopteryx. Экстенсивность заражения рыб достигает 15—20 % при интенсивности инвазии 25—30 гельминтов и более.[ …]
Половозрелая трематода плоская, с длиной тела 1,5 мм и шириной 0,5—0,7 мм. Тело четко разделено перетяжкой и состоит из расширенного переднего и суженного заднего отделов. Хорошо выражены ротовая и брюшная присоски, две ветви кишечника заканчиваются слепо в задней части тела. Яйца овальной формы, размером 0,07—0,09 мм, с крышечкой. Позади брюшной присоски расположен железистый аппарат — орган Брандеса. Размер, форма и строение тела метацер-кария идентичны с таковыми у взрослых гельминтов, но у них недоразвиты половые органы.[ …]
Биология развития. Дефинитивные хозяева — множество видов рыбоядных птиц, в тонком кишечнике которых паразитируют взрослые трематоды. Например, для трематоды A. cornu таковым является серая цапля, Т. intermedia — гагары, Т. diminuta, Т. percaefluviatilis и Т. communis — чайки. Дефинитивными хозяевами указанных выше гельминтов могут быть бакланы и другие рыбоядные птицы. Первыми промежуточными хозяевами являются пресноводные брюхоногие моллюски Bathyomphalus concortus и другие, в теле которых трематоды развиваются партеногенетичес-ким путем. Вторым промежуточным, или дополнительным, хозяином, в теле которого развиваются метацеркарии, являются рыбы различных видов. Например, метацеркарии С. pileatus чаще обнаруживают на серозных покровах полости тела и в плавательном пузыре у ерша, судака, щуки, корюшки и др., Т. percaefluviatilis — на брюшине, стенках плавательного пузыря, сердце, печени, брыжейке, Т. intermedia — на сердце у сиговых и лососевых рыб.[ …]
В 1977-1978 гг. на озерах Игналинской системы Литовской ССР TJL Илюшина /1980/ Ескрыла 1011 ? га. стрекоз семи видов. Из них только у двух найдены метацеркарии трематод: у Aeschna суапеа (4 эка. из 28 вскрытых) – Prosthogonimus ova— tus, ЭИ 14,28 %; у Sympetrum flaveolum (190 из 347 вскрытых) – Prosotocus confusus, ЭИ 62 %. Массовые виды Ischnura elegans (вскрыт 151 экз.), Enallagma cyathigerurr ( 266 экз. ), Erythromma najas (203 экз.) в гельминтологическом отношении оказались стерильными.[ …]
Из Кургальджинских озер Целиноградской области Казахской ССР вскрыто 216 стрекоз восьми видов /Климишин, Павлюк, 1972/. Среди них у пяти видов найдены метацеркарии трематод (ЭИ 22,18): Prosthogonimus sp. – у Aeschna coluberculus, ЭИ 18,18, Sympetrum vulgatum- 10 и S. flaveolum – 0,86; Plagiorchis elegans – у Aeschna juncea (у четырех из шести), Ae. coluberculus — 18,18, S. flaveolum — 15,5, S. scoticum — 24,4, S. vulgatum – 22,66; Prosotocus sp. – у S. flaveolum -5,17. Этот район Д. Кукашев /1979/ отмечает как очаг простогони-моза, где в качестве дополнительных или вторых промежуточных хозяев отмечены стрекозы Libellula quadrimaculata — ЭИ 65 %, Anax parthenope — 63, Aeschna coluberculus ( = Ae.[ …]
На Среднем Урале в Свердловской области HJ3. Чуриной /1964/ вскрыто 364. личинки и 145 имаго стрекоз и установлено, что L. quadrimaculata и Е. najas являются дополнительными хозяевами трематоды Prosthogonimus cuneatus, причем для Е. najas это отмечается впервые. Автором установлен факт развития трематоды Notocotylus attenuatus двумя путями: с образованием адолескарий и с образованием метацеркарий, т.е. с включением дополнительных хозяев, играющих роль резервуара инвазии в водоемах. В условиях Среднего Урала взрослые стрекозы были заражены метацеркариями трематод на 28,2 %, а личинки – на 10,1 %. Личинки больше заражены в естественных прудах (17,9 %), значительно слабее в пойменных (1,3 %), а в озере оказались стерильными. Высокая ин-вазированность (43,1 %) личинками трематод отмечена и у взрослых стрекоз, отловленных вблизи естественных прудов. Стрекозы, пойманные у озера и пойменных прудов, были свободны от инвазии.[ …]
Моллюски и ракообразные, с одной стороны, служат кормом для многих рыб (карповых, лососевых, осетровых, бычков, камбал и др.) и, с другой — являются промежуточными хозяевами паразитических трематод (моллюски) и цестод (веслоногие рачки).[ …]
У S. vulgatum, I. elegans, E. najas, E. cyathigerum, Platycnemis sp. найдены метацеркарии Plagiorchis elegans -ЭИ 6,9 %, ИИ 1—7 экз.[ …]
Довольно редкими паразитами были метацеркарии Halipegus ovocaudatus — ЭИ 0,54 % (2 вида) и Corgodera sp.-0,54 (1 вид). В качестве дополнительных хозяев впервые зарегистрированы: S. meridionale — для трематод Pleurogenoides medians, Prosotocus confusus и Prosthogonimus sp.; C. erythraea — для P. confusus, Skrjabinoeces similis, Plagiorchis elegans и Prostho— gonimus sp.; E. viridulum — для P. confuisus; C. ornatum-для P. confusus и Prosthogonimus sp.; I. elegans – для Prosthogonimus sp.[ …]
В пресных водоемах и болотах зараженность стрекоз личинками трематод достигала 43,7-80 %, в других водоемах она была относительно низка – 2,8-17 %. Метацеркариями плягиорхид (их видовая принадлежность, к сожалению, не указана) были заражены стрекозы из мелководных заливов оз. Иссык-Куль: Lestes dryas – ЭИ 6,6 %, Enallagma cyathigerum- 4,0, Orthetrum cancellatum -5,0; из озера подсобного хозяйства Кот -Сары: L. dryas — 27,5 (плягиорхиды – 15,8 и нотокотилиды – 11,7) и E. cyathigerum — 10,9; из пресноводных озерков: L. dryas – 70,6, Sympetrum flaveolum — 48,0 и S. scoticum – 44,3; из болот: L. dry— as — 71,0 и s. flaveolum — 50,0 %.[ …]
Очень часто наблюдалось сопаразитирование в одном хозяине разных видов, составляющих паразитоценозы всех возможных комбинаций. Исследования показали, что множественные инвазии стрекоз паразитами, в частности, метацеркариями разных видов трематод — явление довольно обычное /Павлюк, 1983а/. При этом соотношения видов при множественных инвазиях колеблются в определенных пределах, что можно проиллюстрировать на двух наиболее сильно поражающихся метацеркариями видах: Sympetrum sanguineum и S. vulgatum.[ …]
В окрестностях Воронежа П.А. Положенцевым и В.П. Негробовым /1958, 1967/ в 1956 г. отловлены и вскрыты 1207 личинок и имйго стрекоз. ЭИ 60 %, С. virgo — 54,8 и L. depressa — 15, с максимальной — ИИ 180 экз. Метацеркарии локализовались в грудной полости.[ …]
В результате многолетних исследований в оз. Б. Миассово установлена паразитофауна для 10 видов рыб. Выявлено многообразие паразитов, принадлежащих к шести классам. Анализ исследований показал, что с 1985 г. по настоящее время ведущее место по видовому разнообразию и по встречаемости в паразите фауне рыб занимают трематоды. Выявлены локальные очаги заражения рыб трематодами и расселение паразитических ракообразных среди рыб в оз. Б. Миассово. Проведенные исследования показали существенную разницу в зараженности паразитами сига, щуки, линя, леща, плотвы (см. табл. 52-57). Сезонные изменения зараженности рыб цестодами во многом определяются характером питания и пищевыми связями рыб в разные сезоны года. В отличие от оз. М. Миассово о паразитах рыб оз. Б. Миассово до последнего времени ничего не было известно. К настоящему времени у рыб оз. Б. Миассово отмечено 66 видов паразитов, а также обнаружено в моллюсках 15 новых для заповедника церка-рий (форм) трематод.[ …]
Т.Г. Никулин /1967, 1969, 1971/ сообщает, что при обследовании 15 озер и одной реки в северной часта Белоруссии у 31 % личинок стрекоз обнаружены личинки гельминтов. При экспериментальном скармливании подопытным утятам личинок стрекоз L. sponsa и S. vdgatum утята заразились эхиностоматидами (Echinostoma revolutum и Hypodaereum conoideum), от А. суапеа – трематодой Catatropis verrucosa из сем.[ …]
Несмотря на изменчивость числа видов между зимней и летней выборками язей, можно выделить большую группу общих видов, которые влияли на видовое богатство и способствовали сходству между сообществами. Однако разная численность одних и тех же видов в летнем и зимнем сообществах делала их сходство низким. В обоих сообществах доминантом был 5. globiporum. Эта трематода имеет широкий круг вторых промежуточных хозяев, в который входят пиявки, турбел-лярии, олигохеты и различные двустворчатые и брюхоногие моллюски (Жохов и др., 1996). Такое разнообразие вторых промежуточных хозяев позволяет с успехом заражаться S. globiporum рыбам любого возраста, начиная с сеголеток. Данное обстоятельство в сочетании с широким спектром питания язя (Ключарева, 1960) делает эту трематоду доминантным видом сообщества. Доминирование одного вида – трематоды Oligogono-tylus manteri – наблюдалось в 4 из 7 популяций цихлиды Cichlasoma urophthalmus в Мексике (Sal-gado-Maldonado, Kennedy, 1997). Поскольку трематода О. manteri – это специфичный паразит цихлидовых рыб и встречалась во всех сообществах, авторы упомянутой статьи делают вывод о том, что главную роль в формировании и сходстве сообществ играет филогенетический фактор.[ …]
По данным мечения, локальные стада гольца (Salveli-nus namaycushi) оз. Супериор (Канада) обособлены друг от друга не только По местам, но и срокам размножения [53]. На основании анализа структуры зон сближенных склеритов на чешуе есть указания о возможности дифференциации локальных группировок нерки в бассейне р. Камчатки [6] и идентификации локальных стад дальневосточных лососей по паразитологическим меткам [17]. Использование трематод в качестве биоиндикатора подтвердило существование локальных группировок малопозвонковой сельди в Белом море [22].[ …]
Однако многие формы водной жизни нежелательны для человека, особенно в случае обработки воды для питьевых целей, и именно этот аспект будет рассмотрен в данной главе. Как животные, так и растительные формы планктона забивают фильтрующую среду (песок, смолы), нарушают работу осветлителей и вызывают вынос осадка. При недостаточной обработке воды планктон может вызвать образование колоний в трубах и резервуарах, а также служить переносчиком патогенных микроорганизмов; железо- и серобактерии принимают участие в коррозионных процессах. Многие болезни, вызываемые бактериями, простейшими (такими, как амебы) или червями (биль-харция, трематода и т. д.), передаются через воду. Все эти виды, для которых вода служит либо местом обитания, либо средством распространения, должны быть удалены из воды при обработке.[ …]
Биология развития. Дефинитивными хозяевами являются рыбоядные птицы, преимущественно чайковые, реже — рыбоядные утки. Промежуточные хозяева — пресноводные моллюски Lymneae stagnalis, Radix ovata и др., дополнительные — рыбы (карп, лещ, окунь, щука, форель, толстолобик и др.). В кишечнике птицы гельминты откладывают яйца, которые при выходе наружу содержат мирацидии. Церкарии покидают организм моллюска и, попав в воду, внедряются через кожный покров в мышцы рыб и затем по кровеносным сосудам попадают в глаза, далее в хрусталик, где вскоре превращаются в метацеркариев. Считают, что метацеркарии в рыбе могут быть жизнеспособными 3—4 года. В кишечнике птиц трематоды достигают половой зрелости через 4—5 сут (рис. 63).[ …]
|
Царство Животные (Zoa). Подцарство Многоклеточные (Metazoa), Надраздел Эуметазои (Eumetazoa), Раздел Двусторонне-симметричные (Bilateria), Тип Плоские черви (Plathelminthes), Класс Трематоды, или Сосальщики (Trematoda = Digenea) Трематоды — паразитический класс плоских червей. Сохраняя все основные признаки типа, они характеризуются специфическими особенностями, которые обусловлены их эндопаразитическим образом жизни. Прежде всего это наличие специальных органов прикрепления — присосок и замена ресничного эпителия лишённым ресничек тегументом. Пищеварительная система трематод, как правило, развита хорошо, но имеется тенденция к её рудиментации и исчезновению. Одновременно с этим приобретает особо важное значение способность трематод к восприятию пищевых веществ извне, через покровы. Как и другие паразитические организмы, трематоды разных поколений производят огромное количество яиц и личинок. Трематодам присуща также тенденция к живорождению, полностью осуществлённая на некоторых этапах их жизненного цикла. Жизненный цикл трематодСам по себе жизненный цикл трематод, связанный с чередованием поколений, совершенно не свойствен свободноживущим плоским червям и возник в ходе эволюции, как следствие перехода этой группы к паразитизму. Жизненный цикл трематод представляет собой одно из самых удивительных явлений живой природы. Он очень сложен, проходит с участием нескольких хозяев, сопровождается чередованием поколений и сменой паразитических и свободноживущих фаз развития. Половозрелая гермафродитная особь — марита паразитирует во внутренних органах позвоночных животных. Откладываемые ею яйца выводятся из организма хозяина во внешнюю среду. Это сложные яйца, под плотной скорлупкой которых находится не только яйцеклетка, но и множество желточных клеток, поставляющих необходимые развивающемуся зародышу питательные вещества. В типичном случае яйцо для завершения своего развития должно попасть в воду, и там из него выходит подвижная личинка – мирацидий. За время короткой жизни эта личинка должна проникнуть в первого промежуточного хозяина — моллюска определённого вида, в котором претерпевает превращение (метаморфоз), связанное с упрощением организации. В результате развивается половозрелая фаза — материнская спороциста. Это очень просто устроенный организм, чаще всего мешковидной формы, способный к размножению путём партеногенеза. Партеногенетические яйца материнской спороцисты, обычно называемые генеративными или зародышевыми клетками, дробятся без оплодотворения и дают начало новому, дочернему поколению — редиям. Последние, покидая материнский организм, активно ползают между органами моллюска хозяина. Обычно они обосновываются в его кровеносных лакунах (кровеносная система моллюсков не замкнута) или в пищеварительной железе. Редии, как и материнские спороцисты, размножаются только партеногенетически, отрождая либо новые поколения редий, либо своеобразных хвостатых церкарий, которые представляют собой личинок гермафродитного поколения трематод (мариты). Редии устроены сложнее, чем материнские спороцисты, так как имеют хорошо развитую пищеварительную систему, представленную мускулистой глоткой и кишечником. Второе партеногенетическое поколение — обязательное звено в жизненном цикле трематод, но у многих высших трематод в материнских спороцистах развиваются не редии, а морфологически иные, более просто устроенные партеногенетические организмы — дочерние спороцисты. Об их соотношении с редиями будет рассказано ниже. Здесь ограничимся лишь сведениями о том, что дочерние спороцисты лишены пищеварительной системы и имеют обычно мешковидную или шарообразную форму тела. Они редко отрождают себе подобных, как правило, давая начало лишь церкариям. Все партениты — партеногенетическое поколение трематод — живородящи. Церкарии представляют собой очень подвижных маленьких личинок, снабжённых специальным органом движения — хвостом. Выйдя из тела родительской особи, они покидают организм своего хозяина-моллюска и попадают в воду. Здесь судьба их может быть различной. Церкарии одних видов довольно быстро оседают на прибрежной траве или на заглатываемых нередко птицами подводных предметах (песчинки, мелкие камешки, пустые раковинки моллюсков и тому пподобном). Они одеваются плотной защитной оболочкой (инцистируются) и превращаются в покоящуюся фазу, называемую адолескарией. Будучи случайно проглочена каким-либо позвоночным животным — окончательным хозяином данного вида трематод, личинка выходит из цисты (эксцистируется) и превращается в мариту. Церкарии других видов для дальнейшего развития должны попасть в организм второго промежуточного хозяина (иногда его называют дополнительным). Чаще всего это головастики, мелкие рыбки, личинки водных насекомых, моллюски, пиявки и другие. В организме второго промежуточного хозяина церкарии обычно инцистируются, переходя в фазу метацеркарий. Если зараженный второй промежуточный хозяин будет съеден позвоночным животным, подходящим для развития метацеркарий данного вида, то личинка выходит из цисты и достигает в окончательном хозяине половой зрелости, превращаясь в гермафродитную особь. Таким образом, в жизненном цикле трематод обязательно участвуют три поколения — два партеногенетических и одно гермафродитное. При этом происходит смена двух разных типов полового размножения: обоеполого, связанного с обязательным оплодотворением яйцеклетки, и партеногенетического, которое осуществляется без оплодотворения. В отличие от метагенеза, при котором половое размножение чередуется с бесполым, чередование поколений, свойственное трематодам, называется гетерогонией. Онтогенез, то есть развитие организма от стадии яйца до половозрелости, у всех трёх поколений трематод проходит различно. Онтогенез первого партеногенетического поколения — материнской спороцисты связан с метаморфозом. Её личинкой является мирацидий, который развивается из оплодотворённого яйца мариты. Второе партеногенетическое поколение — будь то редия или дочерняя спороциста — развивается без метаморфоза. Третье, гермафродитное, поколение опять развивается с метаморфозом, но имеет не одну, а две личинки: церкарию и метацеркарию, или адолескарию. Характерно, что первым промежуточным хозяином трематод всегда служит моллюск. Роль второго промежуточного хозяина могут выполнять самые разные животные — от кишечнополостных и гребневиков до млекопитающих включительно. Окончательный же хозяин — позвоночные. По отношению к первому промежуточному хозяину трематоды проявляют узкую специфичность. Это означает, что материнские спороцисты каждого конкретного вида трематод могут паразитировать в моллюсках лишь одного или нескольких близких видов. Напротив, ко второму промежуточному хозяину трематоды, как правило, очень широко специфичны. Метацеркарии одного вида могут поселяться в животных, относящихся не только к разным видам, но и к разным классам и даже типам. Специфичность, вернее приуроченность к тому или иному виду животных — хозяев, объясняется здесь экологическими причинами. Дело в том, что роль второго промежуточного хозяина конкретного вида трематод выполняют лишь те животные, которые служат пищей окончательному хозяину данного вида. Это важное условие для осуществления жизненного цикла паразита. По образному выражению известного немецкого ученого А. Лооса, окончательный хозяин, выискивая свою «излюбленную» пищу, как бы сам подыскивает и собирает своих паразитов. Разумеется, из этого правила могут быть исключения: известны трематоды, которые и на стадии метацеркарий могут быть узкоспецифичны к своему хозяину. Следует ещё сказать несколько слов о взаимоотношениях между трематодами и моллюсками. Выше говорилось об узкой специфичности трематод к их первому промежуточному хозяину. Обращает на себя внимание и удивительная сбалансированность отпошений между партнерами системы паразит — хозяин. Зараженный моллюск, несмотря на наличие паразитов, может долго сохранять жизнеспособность. Морфология и физиология маритыОбычно когда говорят о трематодах, имеют в виду паразитирующую в позвоночных животных мариту половозрелую фазу гермафродитного поколения. Ограничиться описанием только одной мариты, как это часто делается, было бы неверно. Мы уже знаем, что трематоды характеризуются сложным жизненным циклом, который включает по крайней мере три резко отличающихся друг от друга поколения. Поэтому представить особенности строения и биологии трематод можно, только рассматривая каждое из поколений в отдельности. Наиболее сложно устроена марита, которая изменена паразитическим образом жизни менее других поколений и во многом сходна с организацией остальных плоских червей. С описания её строения и биологии мы начнём наше знакомство с трематодами. Тело мариты по большей части имеет уплощённую листовидную форму и несёт одну или две присоски — органы прикрепления, снабжённые сильно развитыми кольцевыми и радиальными мышцами. Передняя (ротовая) присоска всегда пронизана ротовым отверстием, вторая (брюшная) служит только органом прикрепления. Раньше считалось, что обе присоски связаны с ротовыми отверстиями. Отсюда и возникло название трематод — двуустки (Distoma). Покровы трематод представлены типичным тегументом. Его поверхность, лишённая ресничек, часто несёт переплетающиеся гребни и складки, которые играют существенную роль в питании (смотри ниже). Слои кольцевых и продольных мышц хорошо развиты. Паренхима имеет типичное для плоских червей строение, но, в отличие от паренхимы свободноживущих турбеллярий, выполняет ещё и роль депо гликогена, который служит для трематод основным источником энергии (смотори ниже). Пищеварительная система начинается ротовым отверстием, которое ведёт в округлую мускулистую глотку, играющую роль насоса при заглатывании пищи. За глоткой следует пищевод. Он разделяется на две кишечные ветви, заканчивающиеся слепо. У крупных трематод, например у печёночной двуустки, кишечник обильно разветвлён. Эта особенность строения, как и у крупных турбеллярий, имеет важное значение в распределении продуктов пищеварения по телу червя. Длина ветвей кишечника у марит разных видов неодинакова, что связано со способом их питания. Более примитивные трематоды (Fasciola и другие) активно пожирают кровь и плотные ткани или кишечное содержимое хозяина. Иногда они «хищничают», как, например, Paramphistomum, питаясь крупными инфузориями из рубца жвачных (смотри главу «Простейшие»). У таких трематод пищеварительная система развита хорошо. Напротив, у тех видов, которые лучше приспособлены к паразитическому образу жизни, большее значение приобретает способность воспринимать растворённые органические вещества через тегумент. Давно доказано, что даже Fasciola с её хорошо развитым кишечником воспринимает через покровы глюкозу. Не удивительно, что в ходе эволюции трематод, обитающих в среде, насыщенной продуктами пищеварения хозяина, такой способ питания постепенно становится главенствующим. Одновременно с этим их пищеварительная система утрачивает значение и подвергается редукции. Нервная система устроена по типу ортогона, органы чувств развиты слабо. Выделительная система представлена протонефридиями. Половая система трематод устроена сложно и, как у всех плоских червей, гермафродитна. Мужская половая система состоит чаще всего из двух округлых или лопастных (иногда сильно изрезанных) семенников. От них отходят семепроводы, которые, сливаясь, образуют семенной пузырёк — хранилище, в котором накапливаются зрелые сперматозоиды. Этот пузырёк либо лежит в паренхиме, либо заключён в специальный мешок с мускулистыми стенками — сумку цирруса. В ней же находится и ввёрнутый внутрь совокупительный орган — циррус. Женская половая система представлена одним яичником, округлым или (у крупных форм) разветвлённым, а также яйцеводом, желточниками, оотипом, семеприёмником, тельцем Мелиса, лауреровым каналом и маткой. В этой совокупности слаженно работающих органов собственно половой железой является яичник, в нём формируются яйцеклетки — овоциты. Желточники продуцируют крупные клетки, цитоплазма которых забита гликогеном и содержит множество желточных или скорлуповых гранул. Благодаря им создаётся плотная оболочка сложного яйца. Поставляемые желточниками клетки служат для питания развивающегося зародыша. Центральная часть женской половой системы — оотип. В него открываются или из него выходят все протоки системы. Прежде всего это яйцевод, по которому в оотип поступают яйцеклетки; сюда же открывается проток маленького мешочка — семеприёмника, в котором хранится семя, полученное от другой особи. По специальным протокам в оотип поступают и желточные клетки. Здесь происходит и оплодотворение яйцеклетки, и формирование сложного яйца. Овоцит окружается группой желточных клеток. Содержащиеся в них гранулы выходят наружу и окружают образовавшийся клеточный комплекс; впоследствии они дают начало плотной скорлупке сложного яйца, которое проталкивается в широкую трубку матки. В оотип открываются ещё протоки одноклеточных желёз тельца Мелиса, роль которых до конца не выяснена. Не исключено, что они участвуют в образовании одного из слоёв скорлупки сложного яйца и выделяют секрет, играющий роль смазки при прохождении его в матку. Весь комплекс органов работает, как живой автомат, который каждые 20—30 секунд выталкивает в матку новое сформированное яйцо. Накапливающиеся в оотипе некоторых трематод избытки семени и желточных клеток выбрасываются наружу через короткий проток — лауреров канал, открывающийся на спинной стороне тела червя. Матка служит для выведения яиц наружу. Но у многих видов (отряды Plagiorchiida, Cyclocoelida и другие) в яйцах, ещё находящихся в матке, начинается дробление овоцита, а нередко проходит и всё эмбриональное развитие личинки, так что наружу выходят яйца, в которых заключён вполне готовый мирацидий. Это очень выгодно в биологическом отношении, так как развивающиеся эмбрионы полностью избавлены от неблагоприятных воздействий внешней среды (резкие колебания температуры, высыхание и так далее). В результате процент выживающих яиц паразита значительно возрастает. Однако эмбриогенез требует определённого времени, и развивающиеся яйца задерживаются в петлях матки, а так как поступление новых яиц непрерывно продолжается, матка становится очень длинной и её многочисленные петли занимают иногда до 2/3 тела червя. Сами же яйца по сравнению с теми, развитие которых идёт во внешней среде, становятся мельче. Это связано с тем, что развивающиеся в них зародыши получают все необходимые питательные вещества непосредственно от материнского организма. Поэтому число желточных клеток, содержащихся в сложном яйце, сокращается. Разумеется, у таких видов и сами желточники слабо развиты. Несмотря на то, что половая система мариты гермафродитна, самооплодотворение встречается очень редко, что невыгодно для вида, так как даёт генетически однородное потомство. Поэтому оплодотворение, как правило, перекрёстное. Известны даже случаи появления вторичной раздельнополости марит. Таковы, например, многие представители своеобразных паразитов морских рыб из семейства Didymozoidae. Полностью раздельнополым становится всё семейство Schistosomatidae — паразитов кровеносной системы человека и животных. Более крупный самец носит самку в особом «канале», который образован свёрнутыми в широкую трубку краями его листовидного тела. Мариты трематод могут паразитировать в любых тканях и органах позвоночных животных. В зависимости от условий существования в тех или иных органах хозяина возникают соответствующие изменения строения и физиологии паразита. Так, мариты, поселяющиеся в кровеносных сосудах или в каких-либо узких протоках (например, желчных протоках печени), отличаются длинным, нитевидно вытянутым телом. В то же время многие тканевые паразиты имеют почти шаровидную форму. Тесно связано с местом поселения и строение органов прикрепления мариты. У паразитов кишечника, особенно у видов, обитающих в прямой кишке, откуда они легко могут быть выброшены при физиологических отправлениях хозяина, присоски развиты особенно сильно. Напротив, у трематод, паразитирующих в полости тела (например, у паразитов из семейства Cyclocoelidae), присоски дегенерируют. От локализации нередко зависит и способ питания мариты. Так, тенденция к замене активного питания всасыванием растворённых органических веществ через покровы особенно заметно проявляется у паразитов крови. Мариты позвоночных нередко оказываются в условиях острого недостатка кислорода. Это влияет на характер обмена, который приближается к анаэробному, так как нормальное дыхание в большинстве случаев оказывается невозможным. В разных органах позвоночного животного парциальное давление кислорода может быть совершенно различным. С этим связано то обстоятельство, что трематоды не утратили способности использовать кислород, если он имеется. Поэтому им свойствен не чисто анаэробный (как предполагали раньше), а смешанный тип обмена. В зависимости от того, в каких органах поселяются мариты, в их обмене могут преобладать либо анаэробные, либо аэробные процессы. Те виды, которые паразитируют в трахее или в лёгких хозяина, способны к нормальному дыханию. Мариты, питающиеся кровью, по-видимому, могут использовать кислород, связанный гемоглобином крови хозяина. Наконец, в тканях многих трематод содержится гемоглобин, отличающийся повышенным сродством к кислороду. Это позволяет маритам использовать даже то незначительное количество свободного кислорода, которое имеется в среде их обитания. Трематоды, как и другие эндопаразитические черви, получают необходимую для жизнедеятельности энергию в результате расщепления гликогена, содержащегося в их тканях. Жиры не могут служить для них источником энергии, так как их расщепление возможно только в присутствии свободного кислорода. В тканях паразитических червей заключены огромные запасы гликогена. У свободноживущих родичей трематод таких запасов гликогена никогда не бывает. Прежде всего это обусловлено тем, что свободноживущие организмы широко используют в качестве источника энергии жиры. С другой стороны, анаэробное расщепление гликогена — процесс крайне невыгодный в энергетическом отношении. Если в присутствии кислорода гликоген расщепляется до углекислого газа и воды, то в анаэробных условиях он распадается на крупномолекулярные соединения (главный продукт обмена — молочная кислота), при этом высвобождается в 10 раз меньше энергии, чем в первом случае. Вот почему так велики в теле мариты запасы гликогена, составляющие до 60% её сухой массы. Не следует забывать, что много гликогена расходуется на образование бесчисленных яиц этих червей. Тип Плоские черви В начало страницы |
Определение Fluke от Merriam-Webster
\ ˈFlük \2 : одна из долей хвоста кита.
1 : случайно удачный удар в бильярд или пул.
2 : удача открытие было случайностью. Ее второй чемпионат показывает, что первый не был простой случайностью.
Fluke (Flounder) – определение торговых видов Browne
Доступно онлайн( Paralichthys dentatus ): ДИКАЯ ПОГОНКА
СЕВЕРНАЯ АТЛАНТИКА ФЛУК – это больше, чем просто отличная альтернатива более дорогим видам камбалы, таким как камбала, камбала и палтус – это прекрасный вид, который стоит подавать в сыром виде.Fluke также известен как «летняя камбала» за их склонность оставаться на берегу в летние месяцы. Этих рыб также называют «левоглазыми камбалами»: как и все камбалы, которые становятся обитателями дна, один глаз мигрирует в другую сторону, в результате чего получается рыба «правый глаз» или «левый глаз». Fluke можно отличить от всех других подошв и камбал в Северной Америке, поскольку только он является «левым глазом».
Как и другие камбалы, двуустка имеет внешность Джекилла-и-Хайда с темно-коричневой, серой или зеленоватой спиной и белой слепой стороной или брюшком.Fluke – ловкие охотники, которые используют быстрые движения и полный рот, чтобы поймать ничего не подозревающую добычу, такую как пескарей, кальмаров и различных ракообразных. Способные менять цвет, чтобы сливаться с морским дном, они поистине морские хамелеоны.
Подается в свежем и сыром виде, двуустка идеальна для приготовления сашими (обычно обозначаемого японцами как хираме), особенно севиче или крудо. Приготовленное белое слоеное мясо хорошо выдерживает многие виды приготовления, включая сковороду, гриль, пароварку или духовку.Из-за низкого содержания натуральных масел филе двуустки имеет тенденцию к высыханию, и его обычно подают пашотом или жареным с жидкостью, хотя крепкие соусы могут подавить его сладкий и нежный вкус.
Свежие двуусткиBrowne Trading Co. собирают в прохладных водах у берегов Новой Англии и Средней Атлантики и привозят несколько раз в неделю. Размер рынка обычно составляет от 1 до 4 фунтов, но более крупная рыба, известная как «коврики», обычно может достигать 8 фунтов или больше, а иногда и 18-20 фунтов.
Район вылова: Средняя и Северная Атлантика
Сезонность: Круглый год
Метод улова: Ловлено, поймано тралом
Выход ( Процент филе ): 60%
Профиль вкуса: Мягкий, но сладкий
Профиль текстуры: Lean and Flaky, Firm
Заменитель:
Все, что вам нужно знать
Время чтения: 4 минуты.Камбала – одна из любимых прибрежных рыб Америки.Это весело ловить, вкусно есть, и оно проявляется вдоль Атлантического океана и в Мексиканском заливе. Единственная проблема в том, что существует гораздо больше, чем один вид. Fluke vs. Flounder, Summer vs. Winter, Southern vs. Northern – может быть сложно сказать, какой из них вас зацепил.
Это камбала, но какая?Если вы не знаете, что такое популярная в стране камбала, не бойтесь! Эта статья посвящена разнице между Fluke и Flounder. Вы можете узнать, где они живут и как их отличить.Вы также можете расшифровать различные имена, которые эти вкусные обитатели дна набирали за долгие годы.
В чем разница между Fluke и Flounder?
Проще говоря, Fluke – это Flounder. Fluke – это еще одно название летней камбалы, крупного хищного вида камбалы, обитающего в Северной Атлантике. Причина, по которой люди путаются, заключается в том, что там же живут и зимние камбалы. К счастью, есть несколько простых способов отличить их друг от друга.
Камбала специально приспособлена к жизни на дне.Настолько, что с возрастом один глаз «мигрирует» по их голове, пока оба глаза не окажутся на одной стороне лица. Fluke «обращены влево», что означает, что когда вы поднимаете одного из них, его голова будет в вашей левой руке, когда их глаза будут над их ртом. Зимняя камбала наоборот («направо»).
Это нормально, когда вы держите рыбу, но что, если она в воде? Легко: зимняя камбала темнее, чем летняя камбала. Они могут выглядеть почти черными – отсюда их прозвище «Блэкбэк».Летняя камбала светло-коричневая с кремовыми или белесыми пятнами. Однако камбалы могут менять цвет в соответствии с окружающей средой, поэтому их лица – самый верный способ отличить их друг от друга.
Все становится немного сложнее, когда вы приезжаете в Южную Камбалу. Южная камбала похожа на Fluke. Он такого же размера и цвета, и указывает на то же самое. Так как же их отличить? Просто: пятна. У летней камбалы есть три окольцованных пятна в виде глаз возле хвоста и еще несколько на спине.Вместо них у южной камбалы есть темные пятна, как будто кто-то забыл нанести на них пятна.
Распределение камбалы
Не волнуйтесь, в большинстве случаев вам не придется беспокоиться о различиях между южной и летней камбалой. Причина в том, что они предпочитают разные воды. Имея это в виду, вот посмотрите, где живут разные камбалы. Ключ в названии.
Южные камбалы появляются вдоль Мексиканского залива и вверх по Атлантике до южного Мэриленда.Fluke также известен как «северная камбала». Вы можете найти их в Каролинах, но они гораздо чаще встречаются к северу от Чесапикского залива. Зимняя камбала обитает на крайнем севере, начиная с Мэриленда и заканчивая Канадой.
Fluke vs. Flounder: подводя итоги
На восточном побережье обитают три основных вида камбалы. Вот краткое описание того, что делает каждый из них особенным:
Камбала летняя
- Летняя камбала также известна как «двуустка» или «северная камбала».”
- Его голова смотрит влево, когда вы держите ее в руках.
- Окруженные, похожие на глаза пятна возле хвоста и на спине.
- Обитает от Северной Каролины до штата Мэн.
Камбала зимняя
- Зимняя камбала также известна как «камбала блэкбэк».
- Его голова находится справа, когда вы держите ее.
- Он намного темнее других камбал.
- Обитает от Мэриленда до Лабрадора в Канаде.
Южная камбала
- Южная Камбала повернута «налево» и похожа на Флюка.
- На нем темные пятна вместо кольцевидных, похожих на глаза пятен.
- Предпочитает более теплые воды, чем большинство видов камбал.
- Обитает от Мэриленда до Флориды и в Мексиканском заливе.
Это три основных вида, но камбалы гораздо больше. Надеюсь, нам удалось развеять некоторые недоразумения, связанные с этой восхитительной камбалой. Как и в случае со многими другими вещами на рыбалке, лучший способ изучить остальное – это отправиться в путь и собрать одну из них самостоятельно!
Какая ваша любимая камбала? Вы знаете какие-либо другие способы отличить их друг от друга? Напишите нам в комментариях ниже, мы будем рады услышать от вас!
Fluke
Fluke, также известный как летний камбала, легко узнаваема, потому что она приплюснута из стороны в сторону, позволяя им лежать на песке или илистое дно, частично зарываясь в ожидании ничего не подозревающая рыба-наживка.
Во время личиночной стадии у двуустки правый глаз перемещается к левая сторона, верхняя сторона рыбы. Этот верхний сторона может меняться от светло-коричневой до почти черной, что позволяет рыба, которую нужно смешать, когда она лежит на дне. В правая или нижняя сторона белая, что затрудняет рыбу чтобы увидеть снизу, когда он находится вверху в толще воды.
Fluke известны как прожорливые хищники. У них острый зубы и умеют питаться мелкой рыбой.Большая случайность, известные как “коврики” по понятным причинам, могут подниматься вверх пятнадцать фунтов, но наиболее распространенный размер – от двух до четырех фунты стерлингов.
Мы ловим двуустку в заливе или океане, дрейфуя с копьями. и кальмар. Лучше всего кусает двуустка, когда мы можем дрейфовать примерно на 1 морской узел. Когда мы ловим рыбу в заливе, мы можем ловить рыбу в разных местах в разных частях прилива, чтобы найти правильное количество тока для хорошего дрейфа. При рыбалке в в океане обычно слабое течение, поэтому постарайтесь выбрать день с легким ветерком, чтобы у нас было достаточно дрейфа.От 10 до 20 миля в час ветер обычно самый лучший.
двуустка – одна из самых распространенных рыб в наши воды. Их запасы увеличились в четыре раза больше, чем они были всего 50 лет назад. Несмотря на их обилие Новичкам бывает непросто уловить случайность, поэтому Мы включили несколько советов, которые могут оказаться полезными.
Советы для рыбалки:
Базовый:
Fluke хватает наживку на полпути между хвостом и головой и подержите около 5 секунд, прежде чем они попытаются проглотить Это.Вы должны подождать примерно 10 секунд после того, как почувствуете лишний вес на вашей леске, чтобы начать наматывать. Ошибка многих Новый способ ловли двуустки – поднимать удочку, когда они чувствуют кусать. Теперь попробуйте представить, как ваша леска спускается к грузилу. и трехфутовый лидер почти под прямым углом. Когда ты поднимите грузило на две фута со дна крючок и наживку сдвинуться всего на несколько дюймов, этого недостаточно, чтобы зацепить удачу. Эти два подъем удилища для сибаса, порги и blackfish, потому что мы ловим на двенадцатидюймовый поводок.
Теперь вы подняли жезл на две ноги, опытные рыбаки научились держать удочку вверх и начать наматывать. Это зацепит рыбу. Практически все новички поднимите удочку на две ноги, почувствуйте вес лапы, опустите удилище, а затем начните наматывать. Когда вы опускаете стержень крючок больше не прижимается к внутренней части сосальщики костлявый рот. Счастливчик чувствует, что что-то не так, и открывает рот.Когда вы начинаете наматывать приманку и крючок выйти из его рта. Лучший способ поймать двуустку особенно новичку стоит ждать лишнего веса на твоей леске это двуустка плывет вместе с твоей приманкой на полпути во рту. Примерно через пять или десять секунд он несколько раз перевернет хвост для прибавки скорости, открой его рот и проглотить наживку и крючок. Подождав несколько секунд или почувствовав несколько резких рывков (счастливчик переворачивает хвост и жевание наживки) не двигайте удочку, просто начните шатается.Постоянное давление крючка на лапы пасть ловит рыбу почти каждый раз. Кроме того, если двуустка не на крючке она все еще может поймать и съесть наживку крюк.
Вот несколько советов по рыбалке при дрифте условия не идеальные. При медленном дрейфе сторона лодка с леской, идущей под лодку, лучше всего, потому что эти приманки первыми добираются до рыбы. С быстрым дрейфом сторона, где лески отходят от лодки, лучше всего, потому что как счастливчик пытается поймать наживку и добраться до эти приманки в первую очередь.
Продвинутый:
В дни с небольшим ветром в океане и медленным дрейфом, наиболее эффективна джиггинг при помощи кеглей и ударов. При ловле на океанскую двуустку при сильном ветре и быстром дрейфе плетеная леска обеспечивает меньшее сопротивление, а ваше грузило и наживку лучше оставаться на дне. Если у вас есть сомнения по поводу вашего размер грузила используйте более тяжелое. Ваша линия должна быть на Нижний!
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Инфекции печеночного тремора | Энциклопедия.com
Введение
История болезни, характеристики и передача
Масштабы и распространение
Лечение и профилактика
Воздействие и проблемы
БИБЛИОГРАФИЯ
Инфекции печеночных двуусток являются результатом заражения печеночными червями. Существует два основных типа инфекций, вызываемых печеночными двуустками: фасциолез и фисторхоз. Хотя каждая инфекция вызывается разными видами сосальщиков, у них есть общие черты по характеристикам и передаче.Люди заражаются, когда глотают цисты, содержащие паразитарные формы сосальщиков. Эти кисты открываются в пищеварительной системе и высвобождают паразитов. Люди чаще всего проглатывают цисты после питья зараженной воды или употребления сырой или недоваренной пищи, содержащей цисты.
Инфекция может протекать бессимптомно, быть острой или хронической. Легкие случаи фасциолеза и описторхоза приводят к усталости, лихорадке, болям, опуханию печени, боли в животе и сыпи. Симптомы хронических форм включают обострение острых симптомов с возможной диареей, тошнотой, отеком лица, закупоркой желчных протоков, а иногда и такими осложнениями, как миграция сосальщиков в другие области тела.Обычно эффективным является прием одного из множества противоглистных препаратов, при котором может наступить выздоровление.
Фасциолез встречается во всем мире, тогда как описторхоз обычно встречается в регионах Азии. В Китае зарегистрировано учащение случаев инфицирования печеночной двуусткой, что, как утверждается, является следствием увеличения потребления сырых продуктов.
СЛОВА, КОТОРЫЕ НЕОБХОДИМО ЗНАТЬ
ПРОТИВОГЕЛЬМИНТИЧЕСКИЕ: Противогельминтные препараты – это лекарства, которые избавляют организм от паразитарных червей.
АСИМПТОМАТИЧЕСКИЙ: Состояние, при котором у человека нет симптомов заболевания.
CYST: Относится либо к закрытой полости или мешочку, либо к стадии жизни некоторых паразитов, во время которой они живут в замкнутом пространстве. Этап в жизни простейших, когда они покрыты жесткой внешней оболочкой и впадают в спячку.
ХЕЛЬМИНТ: Представитель различных типов червеобразных животных.
LARVAE: Незрелые формы (червеобразные у насекомых; рыбоподобные у земноводных) организма, способного выживать самостоятельно.Личинки не похожи на своих родителей и должны пройти метаморфоз или измениться, чтобы достичь взрослой стадии.
ПАРАЗИТ: Организм, который живет в организме хозяина или на нем и получает питание от этого хозяина. Паразит обычно получает все преимущества этих отношений, в то время как хозяин может страдать от различных заболеваний и дискомфорта или не проявлять никаких признаков инфекции. Жизненный цикл типичного паразита обычно включает несколько стадий развития и морфологических изменений по мере того, как паразит живет и перемещается через окружающую среду и одного или нескольких хозяев.Паразиты, которые остаются на поверхности тела хозяина для питания, называются эктопаразитами, а те, которые живут внутри тела хозяина, называются эндопаразитами. Паразитизм – это очень успешная биологическая адаптация. Существует больше известных видов паразитов, чем непаразитарных, и паразиты поражают практически все формы жизни, включая почти всех животных, растения и даже бактерии.
ТРЕМАТОДЫ: Обычно известные как сосальщики; класс червей, для которых характерны плоские тела овальной формы.
Инфекции, вызываемые двуусткой печеночной двуусткой, вызывают печеночные двуустки, которые являются разновидностью гельминтов или паразитических червей. Есть два основных заболевания, от которых страдают люди, инфицированные печеночными двуустками: фасциолез, или инфекция печеночного сосальщика, и описторхоз, или инфекция китайского печеночного сосальщика.
Печеночные двуустки принадлежат к особой группе паразитических червей, известных как двуустки или трематоды. Фасциолез вызывается разновидностями двуустки из рода Fasciola , а именно одним видом, Fasciola hepatica , двуусткой овечьей печени.Жизненный цикл этой двуустки выглядит следующим образом: яйца выходят из фекалий хозяина в воду; личинки из яиц заражают улиток; улитки выпускают на растительность личинок, которые образуют цисты, содержащие стадии инфекционного паразита; люди заглатывают цисты, когда едят растения (например, кресс-салат). Затем кисты прорываются в пищеварительном тракте человека, и двуустки проникают в печень и разрушают ткани. Описторхоз вызывается разновидностью двуустки из рода Clonorchis или Opisthrochis , а именно C.sinensis , O. viverrini или O. felineus . Эти двуустки имеют такой же жизненный цикл, что и двуустки из овечьей печени, за исключением того, что они инцистируются не на растениях, а внутри рыбы. Затем люди потребляют рыбу и заражаются.
Инфекции, вызванные двуусткой печеночной двуустки, встречаются во всем мире, хотя распространение фасциолеза и описторхоза различается. Известно, что фасциолез встречается во всем мире как в умеренных, так и в тропических регионах. F. hepatica Сообщалось о инфекциях в Европе, на Ближнем Востоке и в Азии. F. gigantica Сообщалось о случаях заражения в Азии, Африке и на Гавайях. В целом, фасциолез тесно связан с регионами, в которых разводят овец и крупный рогатый скот. Овцы и крупный рогатый скот являются естественными хозяевами печеночных сосальщиков и, следовательно, могут передавать паразита людям, находящимся в тесном контакте с животными или их водоснабжением.
Описторхоз встречается в Азии и Европе. В частности, инфекции O. viverrini зарегистрированы на северо-востоке Таиланда, Лаоса и Кампучии. инфекции, вызванные O. felineus. инфекции были зарегистрированы в основном в Европе и Азии. Почти весь Китай содержит инфекции O. sinensis .
В Китае официальные органы здравоохранения сообщили о 75% -ном увеличении числа случаев печеночной двуустки с 2001 по 2004 год. Причина увеличения связана с повышенным желанием потреблять сырые или недоваренные морепродукты и мясо, которые являются источник печеночных сосальщиков. Как следствие этого увеличения числа инфекций, также увеличилось количество случаев, когда заболевание печени развилось из-за сосальщиков.
Лечение инфекций, вызванных двуусткой печеночной двуусткой, осуществляется с помощью лекарств. Существует ряд эффективных препаратов, включая триклабендазол, празиквантел, битионол, альбендазол и мебендазол. Лечение инфекции, вызванной Fasciola hepatica , обычно включает триклабендазол или битионол. Использование празиквантела в некоторых случаях было неэффективным, и поэтому CDC не рекомендует его использовать для лечения инфекций, вызываемых сосальщиками F. hepatica . Инфекции, вызываемые описторхозом, можно эффективно лечить с помощью празиквантела, который является предпочтительным методом лечения, рекомендованным Центрами по контролю и профилактике заболеваний (CDC).Эти лекарства действуют для уничтожения паразита. Празиквантел действует, парализуя аппарат прикрепления сосальщиков, что лишает их возможности оставаться прикрепленными к кровеносным сосудам хозяина. Это приводит к гибели паразитов, и в конечном итоге инфекция проходит.
Иногда лечение с использованием вышеупомянутых препаратов может вызывать побочные эффекты, такие как диарея, головокружение или головная боль. Однако после лечения вероятно полное выздоровление. В некоторых случаях повреждение печени в результате прикрепления сосальщиков к тканям может сделать пациентов уязвимыми для других инфекций.Лечение может потребоваться в течение нескольких дней или недель в зависимости от типа двуустки, вызвавшей инфекцию.
Вакцины против печеночной двуустки не существует, поэтому предотвращение заражения этими паразитами является лучшим методом профилактики. Избежать этого можно путем кипячения или очистки питьевой воды, тщательного приготовления пресноводной рыбы и растений перед употреблением, а также искоренения или контроля улиток, поскольку они являются промежуточным хозяином для сосальщиков.
В КОНТЕКСТЕ : БОЛЕЗНИ В РАЗВИВАЮЩИХСЯ СТРАНАХС таксономической точки зрения печеночные двуустки относятся к классу Trematoda, который затем подразделяется на два отряда облигатных паразитов: Digenea и Monogena.Облигатные паразиты – это паразиты, которые не могут жить независимо от хозяина. Трематоды, важные для болезней человека, являются частью отряда Digenea и включают кровь, ткани и кишечные сосальщики. Все они в основном встречаются в развивающихся странах, расположенных в тропических и субтропических регионах. Сосальщики редко представляют собой естественную проблему в умеренных климатических зонах.
Печеночные двуустки передаются человеку при употреблении в пищу сырого мяса, рыбы и других растений. Таким образом, пищевые привычки людей имеют сильное влияние на распространенность инфекций, вызываемых печеночными двуустками.В Китае рост потребления сырого или недоваренного мяса и морепродуктов привел к значительному увеличению числа людей, инфицированных печеночными двуустками. В течение 2001–2004 годов китайские чиновники здравоохранения сообщили об увеличении числа случаев инфицирования двуустками на 75%, подчеркнув возможность того, что повышенное потребление пищи, потенциально содержащей печеночных двуусток, вызывает рост инфекции.
Печеночная двуустка F. hepatica , также известная как печеночная двуустка овец, часто поражает домашний скот, например овец, крупный рогатый скот и свиней.Это создает проблемы для производства говядины, баранины и свинины, поскольку сосальщики могут нанести значительный ущерб животным и стать источником инфекции для населения. Профилактика заражения скотом печеночной двуусткой требует регулярного заражения животных гельминтами, а также применения таких методов борьбы с улитками, как исключение животных из районов, зараженных улитками, или осушение водоемов, содержащих улиток. Заболевание может возникать во влажных местах, где обитают улитки. Если улитки отсутствуют, то и эти двуустки из-за их зависимости от улиток тоже.
Заболеваемость печеночными двуустками наиболее высока в районах, где отсутствуют надлежащие средства санитарии и очистки воды. Более эффективные методы санитарии, включая надлежащее удаление и обработку отходов жизнедеятельности человека и домашнего скота, предотвращение загрязнения источников воды фекалиями и более безопасные методы хранения и приготовления пищевых продуктов, могут значительно снизить количество заболеваний, вызываемых всеми гельминтами. Однако Всемирная организация здравоохранения (ВОЗ) отмечает, что более одного миллиарда человек во всем мире не имеют доступа к чистой, незагрязненной воде.В сочетании с диетой, богатой продуктами, которые чаще всего являются переносчиками печеночных сосальщиков, антисанитарные условия затрудняют профилактику печеночных сосальщиков в слаборазвитых регионах.
См. Также Пищевые болезни и безопасность пищевых продуктов; Гельминтозы; Инфекция легочной двуустки (Paragonimus); Оппортунистическая инфекция; Паразитарные болезни.
Веб-сайты
Кембриджский университет . «Fasciola hepatica: печеночная двуустка». 5 октября 1998 г.
Кембриджский университет . «Opisthorchis sinensis: китайская печеночная двуустка». 5 октября 1998 г.
Центры по контролю за заболеваниями (CDC) . «Фасциолез». 6 мая 2004 г.
Центры по контролю за заболеваниями (CDC) .«Описторхоз». 6 мая 2004 г.
Meat Promotion Wales . «Печеночная двуустка».
ProMED-Mail . «Пищевые паразитарные инфекции увеличиваются в Китае». 18 мая 2005 г.
Печеночный Fluke
Печень Fluke
(фасциолез)
Что такое печеночная двуустка?
Печеночная двуустка (фасциолез) вызывается конкретным плоским червем, известным как Fasciola hepatica . У этого паразита сложный жизненный цикл, который обычно занимает от 9 до 20 недель от начала до конца. Незрелые стадии печеночной двуустки также способны выживать в кистообразной структуре, подобной Dictyocaulus. В этом состоянии инцистированная двуустка может выжить до 2 лет, особенно в мягкие зимы.Это имеет косвенный эффект для мер контроля, так как означает, что после того, как случайность установлена, программы контроля должны выполняться из года в год.
Уровни двуустки печеночной двуустки в Британской Ирландии в последние годы неуклонно повышаются, и фермы на всех типах земель, в том числе на традиционно более засушливых землях, теперь должны обеспечить наличие стратегий надзора и контроля над двуусткой. Мягкая погода последних зим в сочетании с обильным количеством осадков в последнее время летом идеально подходит для большого количества двуусток и улиток, и многие скотобойни сообщают о тенденции к увеличению вредного воздействия на печень.
Как влияет печеночная двуустка? Если у моего крупного рогатого скота нет явных симптомов, могут ли они быть случайными?
Даже если у вашего поголовья нет очевидных клинических симптомов, таких как челюсть от бутылочки, диарея и потеря веса, это не означает, что в вашем поголовье нет случайностей.
Субклиническая удача у крупного рогатого скота вызывает снижение прибыльности из-за отсутствия прироста, снижения эффективности преобразования корма, снижения фертильности, повышенной восприимчивости к другим инфекциям и снижения удоя, поэтому важно не предполагать, что у вашего скота нет счастливых случайностей.
Насколько велика проблема?
В результате крайне негативного воздействия на благополучие и продуктивность животных печеночная двуустка представляет собой серьезную экономическую проблему для многих британских животноводов. Такие проблемы, как снижение продуктивности молока, снижение веса и снижение фертильности, неизбежно оборачиваются серьезными расходами для фермера, и исследование 2003 г. показало, что печеночная двуустка обходится животноводческой отрасли Великобритании в 300 миллионов фунтов стерлингов в год. 1 По оценкам того же исследования, двуустка поражает 377 голов крупного рогатого скота на 1000 голов.Более того, британские эксперты прогнозируют, что в ближайшие 60 лет число случаев заражения двуусткой достигнет беспрецедентного уровня. Принимая во внимание такие отрезвляющие цифры, неудивительно, что эффективная борьба с печеночной двуусткой остается главным приоритетом для фермеров и специалистов по ветеринарии в Великобритании.
В результате крайне негативного воздействия на благополучие и продуктивность животных печеночная двуустка представляет собой серьезную экономическую проблему для многих ирландских животноводов. Такие проблемы, как снижение продуктивности молока, снижение прибавки в весе и снижение фертильности, неизбежно приводят к серьезным расходам для фермера.В отчете Animal Health Ireland Report , « Liver Fluke – The Facts » отмечается, что ежегодно печеночная двуустка «оценивается для промышленности Ирландии как минимум в 90 миллионов евро». В отчете также отмечается, что «печеночная двуустка может вызвать сокращение производства мяса до 20% у крупного рогатого скота… и сокращение до 8% у коров». Более того, некоторые эксперты предсказывают, что в ближайшие 60 лет число случаев заражения двуусткой достигнет беспрецедентного уровня. Учитывая такие отрезвляющие цифры, неудивительно, что эффективная борьба с печеночной двуусткой остается главным приоритетом для фермеров и специалистов по ветеринарии Ирландии.
Жизненный цикл печеночной двуустки
На приведенной ниже диаграмме показан жизненный цикл печеночной двуустки.
Ключевые этапы жизненного цикла печеночной двуустки:
- Взрослая печеночная двуустка в печени своего хозяина откладывает яйца, которые переходят на пастбище
- Эти яйца вылупляются во влажных и теплых условиях; производит подвижных личинок (мирацидий), которые затем заражают определенные виды улиток
- Личинки размножаются внутри улитки и развиваются в другую стадию (церкарии), которые выходят из улитки и оседают на пастбище
- Затем они развиваются в высокотолерантную немобильную стадию (метацеркарию), которая может выжить не менее года.После того, как корова съела, они вылупляются и попадают в печень.
1.